LWF aktuell 151
Kalabrische Weißtanne in Bayern
von Muhidin Šeho, Daniel Glas, Johann Geiger und Barbara Fussi
Auf Grund von immer häufiger auftretenden Extremwetterereignissen, Trockenperioden im Sommer und der angespannten Borkenkäfersituation kommt es in vielen Teilen Bayerns zum Ausfall der Fichte. Nach der Meinung vieler Forstexperten kann die Weißtanne auf einigen Standorten in Mischung mit weiteren Baumarten bestimmte Flächenanteile übernehmen.
Bedeutung der Weißtanne
Die Weißtanne nimmt in Bayern nach den aktuellen Ergebnissen der Bundeswaldinventur einen Flächenanteil von 2,8 % ein und konnte seit der letzten Inventur an Waldfläche sogar hinzugewinnen. Als schattentolerante Baumart profitiert sie durch die naturnahe Forstwirtschaft und konnte seit der Bundeswaldinventur 2012 ihre Verjüngungsfläche auf über 42.000 ha verdoppeln. Damit ist die Weißtanne in der Verjüngung deutlich stärker vertreten als im Hauptbestand (LWF 2024). Sowohl die Bayerischen Staatsforsten als auch Privatwaldbesitzer entscheiden sich zunehmend für die Weißtanne als Nadelholzalternative zur Fichte. Die letzten Trockenjahre werfen jedoch die Frage auf, ob die heimischen Herkünfte in Zeiten des Klimawandels tatsächlich die einzige Wahl darstellen und nur noch diese verwendet werden sollten. Gerade auf trockenen Standorten besteht ein deutlicher Bedarf, auch andere klimaplastische Herkünfte aus dem natürlichen Verbreitungsgebiet zu nutzen und so die Bestände an den Klimawandel anzupassen. Vor diesem Hintergrund gibt es Überlegungen, bayerische Wälder durch einen Transfer von Herkünften aus wärmeren und trockeneren Regionen Europas, die sich unter den Bedingungen von Versuchsanbauten bewährt haben, anzureichern.
Für die Identifizierung von weiteren klimaresilienten Saatguterntebeständen der Weißtanne in ihrem natürlichen Verbreitungsgebiet wurde im Projekt sensFORclim als Grundlage für künftige Klimawandel-Anpassungsversuche eine Suchkulisse erstellt. Es wurde gezeigt, wie mit Methoden der Nischenmodellierung auf der Herkunftsebene und Klimaanalogie potenziell interessante Herkunftsregionen identifiziert werden können. Die Modelle zielen auf eine möglichst präzise Abbildung des warmen und trockenen Nischenrandes in wärmeren Regionen Europas ab. Damit konnte bspw. genau die interessante Herkunftsregion der Tanne in Kalabrien identifiziert werden. (Abbildung 1) Das Beispiel der kalabrischen Weißtanne (Herkunft „Gariglione") zeigt, dass die hier dargestellte Methode für die Suche nach klimaplastischen Herkünften tatsächlich Erfolg verspricht.
Diese kalabrischen Herkünfte zeigen in Herkunftsversuchen auf trockenen Standorten in Österreich in einer Lage unter 600 m ü. NN hervorragende Ergebnisse. Auch Anbauten mit kalabrischer Tanne in Dänemark und Bayern zeigen sehr gute Vitalität und Zuwächse auch unter warm-trockenen Bedingungen. Um das Risiko auf verschiedene Herkünfte (heimische und nichtheimische) zu verteilen, konzentriert sich das vorliegende bayerische Projekt auf genetisch vielfältige und klimaplastische Populationen aus Kalabrien.
Bestände im natürlichen Verbreitungsgebiet
Bei einer Bereisung der ausgewählten kalabrischen Saatguterntebestände konnte sich das AWG-Team 2023 von den qualitativ hochwertigen Weißtannen am Monte Gariglione und bei Sera San Bruno selbst überzeugen. Im Kern des Sila-Nationalparks im Zentrum Kalabriens am Monte Gariglione liegt der ausgewählte Erntebestand IT/aal/2/D/CAL/0018.
Der 110 bis 180-jährige Weißtannenbestand erstreckt sich in den Höhenlagen von 1.370 bis 1.680 m ü. NN auf einer Fläche von 450 ha. Der Altbestand besteht zu 60 bis 70 % aus Weißtannen, gemischt mit Schwarzkiefern und Rotbuchen. Im Regenstau des Sila fallen hier je nach Höhenlage 820 bis 1.700 mm Niederschlag pro Jahr, davon 70 % von Oktober bis März. Im Sommer fallen max. 100 mm, bei einer Jahresdurchschnittstemperatur von 8,2 °C bis 12,5 °C. Die tiefsten Temperaturen liegen bei –12 °C, die letzten Spätfröste sind Mitte April.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Möglicher Plusbaum einer über 35 m hohen Weißtanne am Monte Gariglione, Kalabrien. (© M. Šeho)
Der 110 bis 180-jährige Weißtannenbestand erstreckt sich in den Höhenlagen von 1.370 bis 1.680 m ü. NN auf einer Fläche von 450 ha. Der Altbestand besteht zu 60 bis 70 % aus Weißtannen, gemischt mit Schwarzkiefern und Rotbuchen. Im Regenstau des Sila fallen hier je nach Höhenlage 820 bis 1.700 mm Niederschlag pro Jahr, davon 70 % von Oktober bis März. Im Sommer fallen max. 100 mm, bei einer Jahresdurchschnittstemperatur von 8,2 °C bis 12,5 °C. Die tiefsten Temperaturen liegen bei –12 °C, die letzten Spätfröste sind Mitte April.
Der Standort zeichnet sich durch tiefgründig durchwurzelbare Braunerden aus Verwitterung von Glimmerschiefer und Granit mit mittlerer Basen- und Nährstoffversorgung aus. Die Stammqualität der Weißtannen ist gut bis sehr gut. Die Tannen sind sehr vollholzig, geradschaftig und zeigen lange, vitale grüne Kronen. Der BHD liegt bei 55 bis 90 cm, die Mittelhöhe des Bestandes bei 36 bis 38 m (Abbildung 2). Beeindruckend sind die hohen Vorräte von 900 bis 1.150 Efm/ha. Die gesamte Ernteeinheit ist seit 1977 als Generhaltungsbestand ausgewiesen.
 Zoombild vorhanden
Zoombild vorhanden
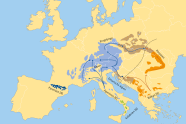
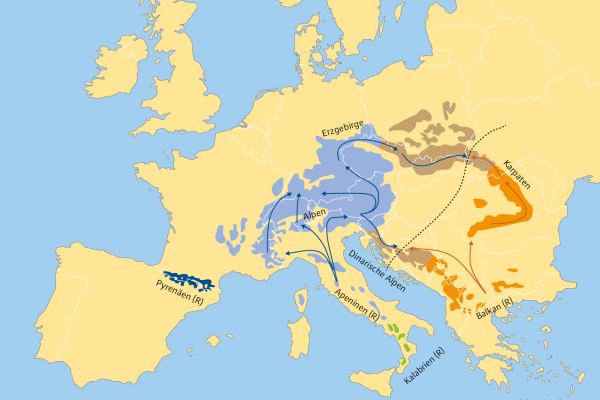
Abb. 4: Phylogenetische Gruppen und Rückwanderungswege der Weißtanne. (© Neophytou 2015)
Auf mehreren Versuchsflächen in Dänemark, Österreich und Bayern (IUFRO Herkunftsversuch 1986/87 und Süddeutscher Herkunftsversuch 1989) zeigt sich die Herkunft vom Monte Gariglione vor allem ab dem Alter 30 als besonders vital und wuchskräftig. Ganz besonders scheint dies für Standorte über 8,5 °C Jahresdurchschnittstemperatur zu gelten; Niederschläge unter 800 mm werden (z. B. Versuchsfläche Tännesberg-Maschen bei Flossenbürg) gut verkraftet.
Nach Neophytou 2015 hat die Weißtanne die letzte Eiszeit in den unterschiedlichen Refugialgebieten überdauert (Abbildung 4). Die Rückwanderung der Weißtanne nach Deutschland erfolgte nach der letzten Eiszeit über unterschiedliche Rückwanderungswege (Ost-/West-Alpen). Die Herausbildung der aktuellen Verbreitungs-Muster in Bayern/Deutschland ist das Ergebnis daraus. Die kalabrische Weißtanne hat es auf dem natürlichen Weg nicht nach Bayern geschafft und ist gerade wegen ihrer hohen genetischen Vielfalt und Diversität interessant im Klimawandel.
Genetische Methoden und Analysen
Bei der Bereisung der ausgewählten kalabrischen Saatguterntebestände wurden Nadelproben gewonnen und für weitere Untersuchungen nach Teisendorf gebracht. Für die genetischen Analysen wurden folgende neun SSR-Marker ausgewählt: NFF7, NFH3, NFF3, NFH15 (Hansen et al. 2005), SF78, SF1, SFb4 (Cremer et al. 2006), Aat05, Aat10 (Postolache et al. 2014).
Für die Beschreibung der genetischen Variation wurden ausgehend von den für die Einzelbäume bestimmten Multilocus-Genotypen zuerst die Allelhäufigkeiten berechnet. Nachfolgende genetische Indizes beschreiben die genetische Variation innerhalb der Vorkommen und wurden mit dem Programm GenAIEx Version 6.5 (Peakall & Smouse 2012) berechnet (Abbildung 3).
Mithilfe eines Clusterverfahrens (Structure 2.3.4, Pritchard et al. 2000) wurden mögliche genetische Unterschiede untersucht und graphisch dargestellt (Clumpak (Kopelman et al. 2015)).
| Genetische Variation | Index | Beschreibung |
|---|
| Genetische Vielfalt | Na | durchschnittliche Anzahl der Allele pro Genort |
| Np | Private Allele, die nur in dieser Population vorkommen |
| Genetische Diversität | Ne | mittlere effektive Anzahl von Allelen pro Genort |
| Heterozygotiegrad | Ho | Anteil beobachteter mischerbiger Individuen |
| He | Anteil erwarteter mischerbiger Individuen |
| Fixierungskoeffizient | F | Verhältnis zwischen (Ho) und (He), das den Einfluss von Selektion,
Populationsgröße, Inzucht und Genfluss auf eine Population wiedergibt
|
Abb. 3: Populationsgenetische Parameter zur Identifizierung der genetischen Vielfalt in Saatgutbeständen.
Populationsgenetische Parameter
 Zoombild vorhanden
Zoombild vorhanden
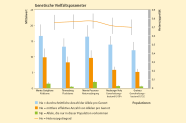
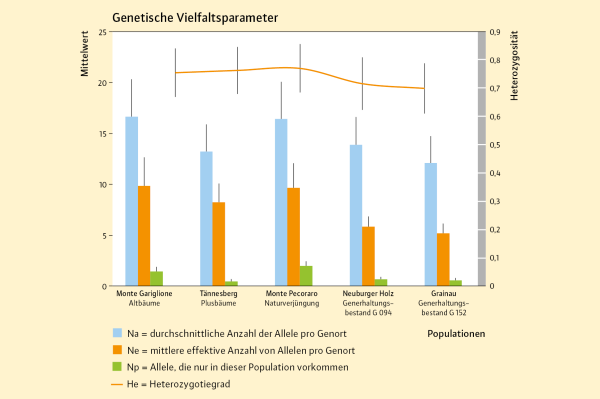
Abb. 5: Genetische Vielfalts-parameter kalabrischer und bayerischer Herkünfte: Monte Gariglione und Monte Pecoraro in Kalabrien, sowie Tännesberg, Neuburger Holz und Grainau in Bayern. (© LWF)
Für die Anpassungsfähigkeit einer Herkunft in der Zukunft ist es wichtig, dass die Ausgangsbestände, in denen Saatgut oder Edelreiser für die Anlage von Samenplantagen gewonnen werden, eine hohe genetische Vielfalt ausweisen. Die vier Tannenbestände sowie die 40 Plusbäume wiesen eine relativ hohe genetische Variation auf. Die Anzahl der Allele (Na) schwankte zwischen 12,1 (Generhaltungsbestand Grainau) und 16,6 (Monte Gariglione) mit einem Gesamtdurchschnitt von Na = 14,5. Die Anzahl der effektiven Allele (Ne) schwankte von 5,2 (Generhaltungsbestand Grainau) bis 9,8 (Monte Gariglione) mit einem Gesamtmittelwert von Ne = 7,7. Diese Werte zeigen die Unterschiede in der genetischen Variation in den untersuchten Populationen. Höhere Werte können eine höhere Anpassungsfähigkeit bedeuten. Die höchste Anzahl an privaten Allelen wurde in der Herkunft Monte Pecoraro (Np = 2,00) gefunden, der geringste Wert lag bei 0,56 (Tännesberg) (Abbildung 5). Private Allele sagen etwas über die Einzigartigkeit der untersuchten Bestände aus. Wenn viele Genvarianten ausschließlich in einer Population vorkommen, ist diese etwas Besonderes und kann daher besonders gut oder besonders schlecht an die Zukunft angepasst sein.
Die beobachtete Heterozygotie (Ho) war in allen untersuchten Populationen ebenfalls unterschiedlich hoch und schwankte zwischen 0,701 (Generhaltungsbestand Grainau) und 0,783 (Tännesberg) bei einem Mittelwert von 0,741. Die erwartete Heterozygotie (He) war in allen untersuchten Populationen ebenfalls unterschiedlich hoch und schwankte zwischen 0,700 (Generhaltungsbestand Grainau) und 0,771 (Monte Pecoraro) bei einem Mittelwert von 0,741 (Abbildung 5). Heterozygote Individuen haben zwei unterschiedliche Genvarianten an einem Genort. Das ist wie ein doppelter Boden. Falls das eine Allel nicht funktioniert, könnte das andere Allel eine Variation tragen, das mit den auftretenden Bedingungen besser zurecht kommt.
 Zoombild vorhanden
Zoombild vorhanden
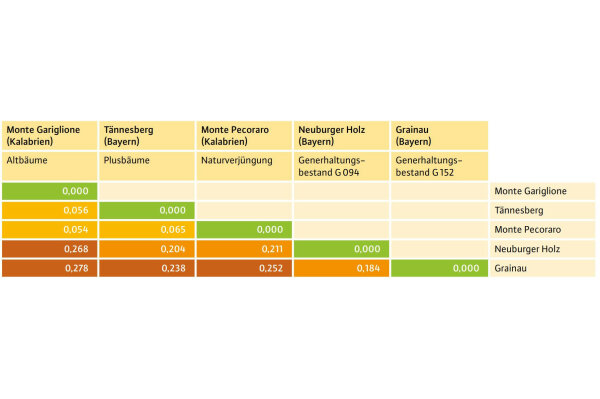
Abb. 6: Genetische Distanzmatrix der untersuchten Tannenbestände nach Nei (1972). (© LWF)
Der beobachtete (Ho) und der erwartete (He) Heterozygotiegrad liegt über alle untersuchten Populationen auf einem ähnlichen Niveau, daher lag der Fixierungsindex (F) nahe Null (–0,4 in Tännesberg und 0,06 in Monte Pecoraro. Es ist also keine Inzucht in den Beständen gefunden worden. Das bedeutet, dass der Genfluss innerhalb und zwischen den Beständen intakt ist.
Die genetische Distanzmatrix nach Nei (1972) (Abbildung 6) zeigt große genetische Abstände zwischen den kalabrischen und den bayerischen Herkünften (0,20–0,28). Die genetischen Abstände zwischen den kalabrischen Populationen sind dagegen sehr gering (0,05–0,06). Auch die im Herkunftsversuch Tännesberg von der kalabrischen Herkunft gewonnenen Plusbäume passen sehr gut zu den aktuell in Kalabrien gesammelten Tannenproben. Das bedeutet, dass die Herkunft „Kalabrische Tanne" im Herkunftsversuch mit sehr hoher Wahrscheinlichkeit aus Kalabrien stammt und die verwendeten genetischen Marker gut geeignet sind, um Kalabrische Tannen zu identifizieren.
Räumlich-genetische Struktur
 Zoombild vorhanden
Zoombild vorhanden
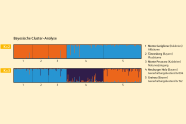
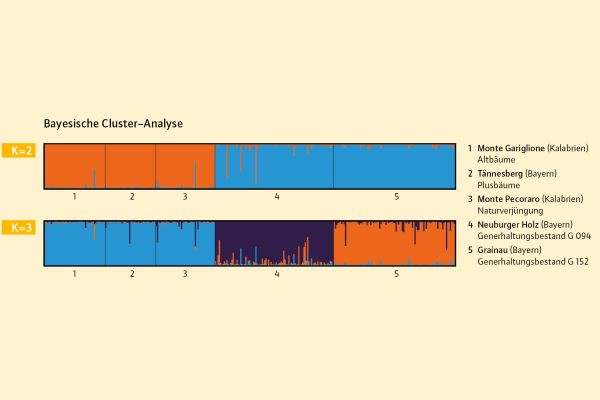
Abb. 7: Genetische Struktur bei zwei Clustern (K=2) und bei drei Clustern (K=3). (© LWF)
Die Ergebnisse der Bayesischen Cluster-Analyse zeigen die beste Gruppierung bei zwei (K=2) und drei Genpools (K=3) an (Abbildung 7). Bei zwei angenommenen Clustern (K=2) liegt der deltaK-Wert bei 166 und zeigt somit eine starke Abgrenzung der kalabrischen von den bayerischen Tannenbeständen an. Eine weitere Gruppierung ergibt sich bei K=3, da die beiden bayerischen Bestände ebenfalls deutlich unterschiedlich sind. Das bedeutet, dass die Plusbäume, die in Bayern auf Versuchsflächen stocken, eindeutig dem kalabrischen Genpool zugeordnet werden können.
Vorgehen in Bayern und Neuanlage der Samenplantage
 Zoombild vorhanden
Zoombild vorhanden
Abb. 8: Vorratsreicher Tannen-bestand am Monte Gariglione in Kalabrien. (© J. Geiger)
Die Kollegen Dr. Schüler und Dr. Konrad vom Bundesforschungszentrum für Wald (BFW) in Wien haben in den letzten Jahren die Gewinnung von Klonen aus Kalabrien detailliert geplant und viele der erforderlichen Arbeiten bereits durchgeführt. Für Bayern bestand nun die Möglichkeit, sich mit einem eigenen Projekt anzuschließen und eine bayerische Samenplantage zu begründen, um langfristig die Versorgung mit trockenheitstolerantem Saatgut der kalabrischen Tanne zu sichern. Als bayerischer Beitrag konnten Reiser von 40 Plusbäumen der kalabrischen Tannen auf verschiedenen Versuchsflächen des AWG u. a. bei Tännesberg identifiziert und gewonnen werden. Die Reiser werden jeweils für eine österreichische und eine bayerische Samenplantage verwendet. Dadurch wird zur Risikostreuung beigetragen und sichergestellt, dass auch bei einem Ausfall von Klonen auf einer Samenplantage eine Kopie auf der anderen Samenplantage gewonnen werden kann. Während der Projektlaufzeit konnten alle geplanten Arbeiten durchgeführt werden. Neben der Plusbaumauswahl, Reisergewinnung und der Veredelung der Bäume konnte eine geeignete Fläche im bayerischen Staatswald bei Stöttwang identifiziert werden. In Zusammenarbeit mit Kollegen vom Forstbetrieb Landsberg am Lech wurde die Fläche geräumt und vorbereitet, so dass die Anlage erfolgen kann.
Zusammenfassung
Die Weißtanne ist eine der wichtigsten heimischen Nadelbaumarten. Gerade auf trockenen Standorten besteht ein deutlicher Bedarf, auch andere klimaresiliente Herkünfte aus dem natürlichen Verbreitungsgebiet zu nutzen und an den Klimawandel anzupassen. Im Rahmen dieser Arbeit wurde die kalabrische Tanne als klimaresiliente Herkunft detailliert untersucht. Für die Anpassungsfähigkeit einer Herkunft in der Zukunft ist es wichtig, dass die Ausgangsbestände eine hohe genetische Vielfalt aufweisen. Die untersuchten Tannen aus Kalabrien weisen eine höhere genetische Vielfalt auf, als die untersuchten bayerischen Tannen aus Generhaltungsbeständen. Die genetischen Unterschiede zwischen den kalabrischen und bayerischen Tannen sind vergleichsweise groß. Um die Versorgung mit genetisch vielfältigem Vermehrungsgut der kalabrischen Tanne zu sichern, werden in Österreich und Bayern Samenplantagen mit hochwertigen Plusbäumen begründet.
Literatur
- Cremer, E., Liepelt, S., Sebastiani, F., Buonamici, A., Michalczyk, I.M., Ziegenhagen, B., Vendramin, G.G. (2006). Identification andcharacterization of nuclear microsatellite loci in Abies alba Mill.Mol Ecol Notes 6(2): 374-376.
- Postolache, D., Leonarduzzi, C., Piotti, A., Spanu, I., Roig, A., Fady, B., ... & Vendramin, G. G. (2014). Transcriptome versus genomic microsatellite markers: highly informative multiplexes for genotyping Abies alba Mill. and congeneric species. Plant Molecular Biology Reporter, 32, 750-760.
- Evanno, G., Regnaut, S., Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, S. 2611-2620. https://doi.org/10.1111/j.1365-294X.2005.02553.x
- Hansen, O.K., Vendramin, G.G., Sebastiani, F., Edwards, K.J., (2005). Development of microsatellite markers in Abies nordmanniana(Stev.) Spach and cross-species amplification in the Abies genus.Mol Ecol Notes 5(4): 784–787.
- KOPELMAN, N. M.; MAYZEL, J.; JAKOBSSON, M.; ROSENBERG, N. A.; MAYROSE, I. (2015): Clumpak: a program for identifying clustering modes and packaging population structure inferences across K. Molecular Ecology Resources 15(5), 1179-1191.
- LI Y. L.; LIU J. X. (2018): StructureSelector: A web based software to select and visualize the optimal number of clusters using multiple methods. Molecular Ecology Resources 18: 176–177.
- NEI, M. (1972): Genetic distance between populations. American Naturalist 106, 283-392.
- Peakall, R., Smouse P. E. (2012). GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28, S. 2537-2539.
- PRITCHARD, J. K.; STEPHENS, M.; DONNELLY, P. (2000): Inference of population structure using Multilocus genotype data. Genetics, 155(2), S. 945 – 959.
- PUECHMAILLE, S. J. (2016): The program structure does not reliably recover the correct population structure when sampling is uneven: subsampling and new estimators alleviate the problem. Molecular Ecology Resources 16: 608-627.
- NEOPHYTOU, C. (2015): Genetik der Weißtanne: Ein- oder Vielfalt?. standort.wald 49 (2015), 85–100.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren



 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden