


Tobias Mette und Wolfgang Falk
Extreme Trockenheit – wie sie auf Vitalität und Anbaurisiko von Waldbäumen wirkt – LWF aktuell 126
Was passiert, wenn Witterungsextreme den Toleranzbereich von Waldbäumen überschreiten?
Jede Baumart hat ihren eigenen klimatischen »Wohlfühlbereich«. Diese sogenannte Klima-Nische wird in der Regel über 20- bis 30-jährige Mittel von Temperatur- und Niederschlagswerten definiert. Sowohl die allseits bekannten Klimahüllen als auch die Artverbreitungsmodelle des Bayerischen Standortinformationssystem BaSIS verwenden ein solches mittleres Klima.
Innerhalb dieser Klima-Nische toleriert eine Art auch gelegentlich auftretende Witterungsextreme wie Spätfröste, Hitze- und Trockenperioden. Im Klimawandel ändern sich sowohl die Mittelwerte – das Klima – als auch die Witterungsextreme. Dabei können Extremjahre wie 2018 oder 2019 den Toleranzbereich einer Art so überschreiten, dass es zu einem regional gehäuften Absterben kommt und die Anzeichen einer Arealverschiebung schlagartig deutlich werden.
Neben 1961–1990 ist der Zeitraum 1971– 2000 eine wichtige Referenzperiode in der Klimawandel-Diskussion. Auch das Anbaurisiko »2000« im Bayerischen Standortinformationssystem – mittlerweile für 32 Baumarten – bezieht sich auf die Periode 1971–2000.
Heute allerdings schreiben wir das Jahr 2020, und im Vergleich zu 1971–2000 ist in der letzten Dekade (2011–2019) die Sommertemperatur bayernweit bereits um 1.6 °C gestiegen, während die Sommerniederschläge um circa 10 % abgenommen haben. Die Gefahr von Trockenstress teils auch in Verbindung mit Hitze hat in den Wäldern Bayerns deutlich zugenommen.
»Hotspots« in Bayern
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Seit 1983 inspizieren jährlich Försterinnen und Förster im Rahmen der Waldzustandserhebung viele tausend Baumkronen. (Foto: T. Hase)
In den Trockensommern 2015, 2018 und 2019 fielen in Mittel- und Unterfranken von Juni bis August durchschnittlich nur 135 mm Niederschlag mit einem Negativrekord von 110 mm in 2018. Die Folgen sind deutlich spürbar: Während die Fichte als Risikobaumart bekannt ist, bereiteten die Ausfälle von Kiefer und sogar Buche vermehrt Grund zur Sorge. LWF aktuell hat hierzu regelmäßig berichtet, zuletzt mit der Ausgabe »Wald unter Druck« (LWF 2020).
Die Reaktion der Bäume und Wälder auf Trockenheit und Hitze wird sehr intensiv erforscht (Schuldt et al. 2020). Zahlreiche Experimente und Feldstudien schärfen das Verständnis für die Physiologie des Baums unter Trockenstress und beleuchten das komplexe Zusammenspiel mit der Bestandeskonkurrenz, mit Boden und Mykorrhiza, und die Rolle von Schadorganismen.
Auch die Bayerische Landesanstalt für Wald und Forstwirtschaft (LWF) beteiligt sich an dieser Forschung, unter anderem zu den Ursachen der Kiefernschäden, zu verbesserten Monitoringmöglichkeiten über Drohnen-, Luft- und Satellitenbilder, oder zu statistischen Auswertungen dürrebedingter Mortalität in Europa (Klemmt et al. 2018; Ackermann et al. 2018; Brandl & Falk 2019).
Waldzustandserhebung und Artverbreitungsmodelle
Zum andern handelt es sich um Artverbreitungsmodelle. Sie stellen einen statistischen Zusammenhang zwischen Klima und Artvorkommen her und sind eine wichtige Grundlage des Anbaurisikos im Bayerischen Standortinformationssystem BaSIS. Allerdings betrachten wir hier ausschließlich die klimatische Komponente und verwenden einen neuen Datensatz europäischer Waldinventuren (Mauri et al. 2017).
Im Fokus der Auswertung stehen 62 permanente Inventurpunkte der Waldzustandserhebung in Mittel- und Unterfranken, die trotz einer Rasterverschiebung im Jahr 2006 eine konsistente Datenreihe von 1994 bis 2019 ergeben. Insgesamt fließen 7.358 Buchen-, 7.682 Fichten- und 17.929 Kieferndatensätze in die Auswertung ein.
Trockenheit und Kronenverlichtung
 Zoombild vorhanden
Zoombild vorhanden
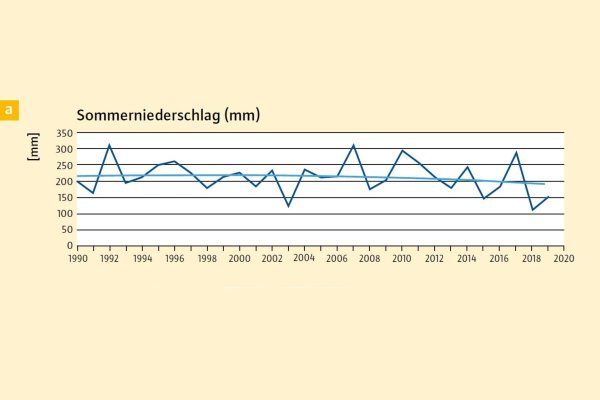
Abb. 2: Sommerniederschlag an 62 ausgewählten Inventurpunkten in Mittel- und Unterfranken (Wuchsgebiete 1 bis 5) (Grafik: LWF)
Sie zeigen einen markanten Nadel-/ Blattverlust der Baumarten Buche, Fichte und Kiefer nach den Trockensommern 2003, 2015 und 2018. Der Verlust erfolgt im Jahr nach dem Trockensommer und beträgt durchschnittlich 5 bis 10 %. Grund für die Verzögerung der Reaktion um ein Jahr ist, dass Blattzahl und Trieblänge maßgeblich von der Entwicklung der Blattknospen im Vorjahr bestimmt werden (Roloff 2001).
Was die WZE-Aufnahme nicht registriert – da sie bereits im Juli abgeschlossen ist – ist, wie die Bäume auch noch im selben Jahr auf einen Trockensommer reagieren: entweder mit vorzeitigem Blattwurf bei anhaltender Trockenheit oder mit verspätetem Blattwurf bei einem warm-feuchten Herbst (Raspe et al. 2004).
Aus Abbildungen 2 bis 4 geht auch hervor, dass die Sommerniederschläge allein zu kurz greifen, um regional gemittelte Blatt- oder Nadelverluste zu erklären. Zum Beispiel hat keine der drei Baumarten auf den trockenen Sommer 1998 reagiert, auf 2008 reagierten nur Buche und Fichte, auf 2013 nur Buche und Kiefer.
Zum einen wirken sich neben Trockenheit und ggf. Hitze auch Faktoren wie Insektenfraß, Fruktifikation oder eine sturmbedingte Erhöhung der Borkenkäferpopulation auf die Laub- bzw. Nadelmasse der Bäume aus. Zum anderen ist die Betrachtung lediglich des Niederschlags zur Beschreibung von Trockenperioden eine sehr starke Vereinfachung des komplexen Wasserhaushalts, der mit entsprechenden Modellen und Bodendaten wesentlich genauer betrachtet werden sollte (Weis et al. in diesem Heft).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Mittlere Benadelung und Belaubung an 62 ausgewählten Inventurpunkten in Mittel- und Unterfranken (Wuchsgebiete 1 bis 5) (Grafik: LWF)
Dadurch erfolgt eine konstante Korrektur zum Gesunden hin. Dieser Effekt wurde von Kölling und Schmidt (2013) als »Lüge der Überlebenden« beschrieben. Zurzeit sieht es allerdings aus, als könnte dieser Effekt die zunehmende Kronenverlichtung von Fichte und Kiefer nicht kompensieren (siehe Abbildungen 2 bis 4).
Auch ein Baumartenwechsel auf trockenheitsgefährdeten Standorten, bei dem vor allem risikoträchtige Arten – zumeist Fichte oder Kiefer – ersetzt werden, bewirkt langfristig eine rein statistische »Gesundung« des Probekollektivs. Zwischen der zweiten und dritten Bundeswaldinventur (2002 und 2012) nahm in Mittel- und Unterfranken der Anteil von Fichte, Kiefer und Lärche zusammen um 4,2 % auf 55 % ab, während der Anteil von Buche und Eiche um insgesamt 1,8 % auf 31 % zunahm. Für die vierte Bundeswaldinventur 2021/2022 ist sicherlich mit noch größeren Änderungen zu rechnen.
Neben der Kronenverlichtung wird im Rahmen der WZE auch das eher seltene Ereignis eines Absterbens aufgenommen. Die zum Teil dürrebedingte Erhöhung der Mortalität im Jahr 2019 ist sowohl im bayerischen (StMELF 2019) als auch im bundesweiten (BMEL 2020) Datensatz der Waldzustandserhebung zu finden und untermauert den Vitalitätsverlust durch Extrema.
Extreme Trockenheit und Anbaurisiko
 Zoombild vorhanden
Zoombild vorhanden
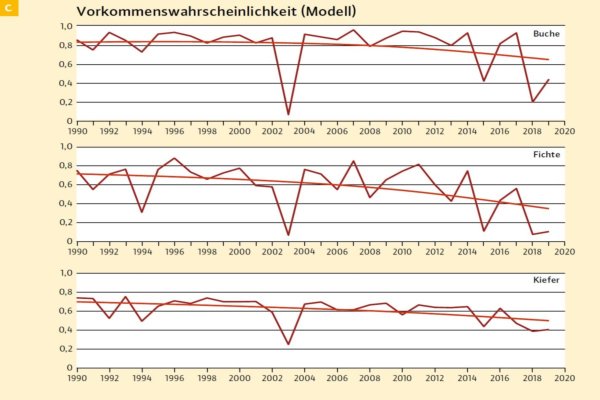
Abb. 4: Vorkommenswahrscheinlichkeit aus Artverbreitungsmodellen an 62 ausgewählten Inventurpunkten
in Mittel- und Unterfranken (Wuchsgebiete 1 bis 5) (Grafik: LWF)
Die hier verwendeten Artverbreitungsmodelle erfassen neben Sommerniederschlag auch Sommer- und Wintertemperatur, um die Vorkommenswahrscheinlichkeit einer Art als Funktion des Klimas zu beschreiben [techn. Anm.: während für Sommertemperatur und -niederschlag die Werte der Einzeljahre verwendet wurden, flossen für die Wintertemperatur geglättete Werte ein. Damit sind die jährlichen Schwankungen primär auf Sommerhitze und -trockenheit zurückzuführen].
Die Wahrscheinlichkeit skaliert zwischen 0 (kommt gar nicht vor) und 1 (kommt zu 100 % vor) und gilt als Indikator für das Anbaurisiko. Da die hier gezeigten Modelle auf anderen Daten beruhen als im Bayerischen Standortinformationssystem, bewerten wir konservativ erst Werte, die unter 0,3 fallen, als kritisch.
Arealverschiebungen: die Folgen häufiger Extremjahre
 Zoombild vorhanden
Zoombild vorhanden
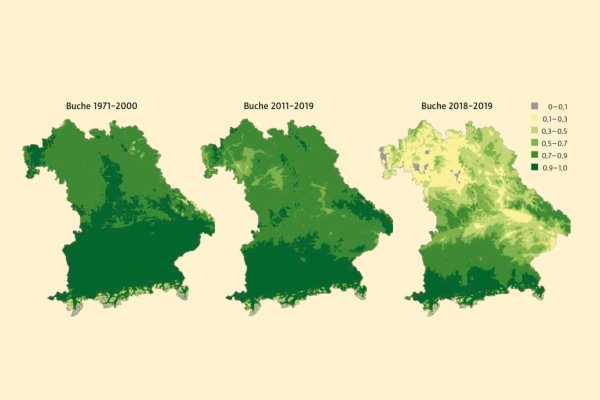
Abb. 5: Wahrscheinlichkeit des Vorkommens der Buche für drei Perioden. Kritische Bereiche sind grau und gelb hinterlegt. (Grafik: LWF)
Auch Buche reagiert stark auf die Trockenjahre, bleibt aber auch im langjährigen Mittel über 0,6. Kiefer reagiert ausgeglichener, was daran liegt, dass das Modell auch durch die Vorkommen in trockenwarmen Gebieten geprägt ist. Trotzdem fällt das Niveau der gemittelten Kurve auf 0,5, und zeigt, dass es sich prinzipiell um eine boreal-kontinentale Baumart handelt. Streng genommen ist eine Anwendung von Artverbreitungsmodellen, die an langfristigen Klima-Mitteln skaliert wurden, auf Einzeljahre nicht korrekt.
Eine jährliche schwankende Vorkommenswahrscheinlichkeit für langlebige Organismen wie Bäume bleibt ein rein theoretischer Wert. Trotzdem eignet sie sich als Indikator für den klimatischen Stress von Baumarten in Trockenjahren, da das Modell einen Bezug zwischen einem Klimaindex und der Verbreitung der Baumart herstellt. Mit einzelnen Extremjahren kann ein Baum in der Regel gut umgehen. Aber wenn solche Extremjahre zur Normalität werden, dann ist die Art auf Dauer nicht überlebensfähig. Dann ist das Vorkommen rückläufig und zeigt eine Arealverschiebung der Art an. 2018/2019 haben deutlich gemacht, dass diese Arealverschiebung nicht allmählich vonstattengeht, sondern sich schlagartig in Extremjahren realisiert.
Dass die Dürren 2018 und 2019 in Teilen Bayerns den Wohlfühlbereich einiger Baumarten verlassen haben, verdeutlicht Abbildung 5 mit einer Gegenüberstellung des Anbaurisikos für Buche in den Perioden 1971–2000, 2011–2019 und für die Ausnahmejahre 2018/2019. Schon beim Vergleich von 1971–2000 mit 2011–2019 zeichnen sich die Tallagen von Regnitz, mittlerem Main und Regen durch eine geringere Vorkommenswahrscheinlichkeit der Buche aus.
Damit tragen die Modelle der deutlichen Klimaverschiebung in den letzten 40 Jahren Rechnung. Würden allerdings Sommer wie 2018/19 langfristig zu den mittleren Sommern zählen, könnte es für den Anbau der Buche in Mittelund Unterfranken sowie im Oberpfälzer Becken und im Donautal kritisch werden. Lange wurden Klimaänderungen solchen Ausmaßes als äußerst unwahrscheinlich angesehen. Mittlerweile zieht man aber auch solche harten Annahmen, dass Sommer wie 2018/19 in einigen Dekaden zur Normalität gehören könnten, in Erwägung.
Extreme Trockenheit und Mortalität
 Zoombild vorhanden
Zoombild vorhanden
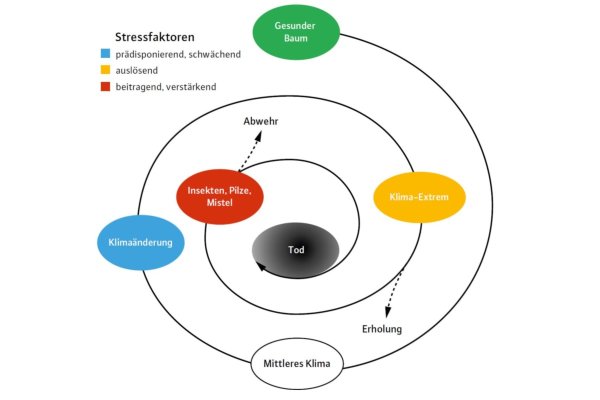
Abb. 6: Mortalitätsspirale nach Manion (1981) angepasst an den Klimawandel-bedingten Trockenstress der letzten Sommer. (Grafik: LWF)
Sehr gut geeignet, um die Komplexität der Mortalität zu verstehen, ist die »Mortalitätsspirale « nach Manion (1981). Sie macht klar, dass es selten nur ein einzelner Faktor, sondern ein Faktorenkomplex ist, der zum Absterben eines Baums führt. In Abbildung 4 haben wir Manions Spirale auf den Baum im Klimawandel übertragen: Wir gehen zunächst davon aus, dass der gesunde Baum unter optimalen Klimabedingungen wächst.
Der Klimawandel bringt Veränderung in Form von immer häufiger auftretenden Extremen, die an einem Punkt den Baum so kritisch schwächen, dass er dem (nicht selten auch witterungsbedingt erhöhten) Schädlingsdruck nicht Stand halten kann. In der Spirale kann man die Klimaänderung als »prädisponierenden« oder »schwächenden« Faktor, das klimatische Extrem als »auslösenden« Faktor und die biotischen Schädlinge als »verstärkende « oder »beitragende« Faktoren betrachten. Weiterhin zeigt Abbildung 6, dass es auch Erholungen oder Abwehrreaktionen gibt, die Abwärtsspirale also auch verlassen werden kann, wenn ein Stressfaktor nachlässt.
Der direkte Dürretod eines Baums – quasi ein physiologischer Kollaps – war in unsern Wäldern bis 2018/19 nur selten zu beobachten. Er ist für die Wissenschaft eine große Herausforderung und es werden zwei Prozesse diskutiert (Mc- Dowell et al. 2008): Zum einen das Verhungern (»carbon starvation«) und zum anderen das Verdursten infolge eines weitgehenden Zusammenbruch der Wasserleitfähigkeit durch Kavitationen, d. h. den Lufteintritt in die Leitungsbahnen (»hydraulic failure«).
Um dies zu verstehen, muss man sich vergegenwärtigen, dass das prinzipielle Dilemma eines jeden Baums darin besteht, dass er, um CO2 für die Photosynthese aufzunehmen, die Spaltöffnungen öffnen muss und dabei automatisch Wasser verliert. Offensichtlich können jedoch einige Arten, zum Beispiel Baumarten mediterraner Regionen, wesentlich extremeren Wassermangel aushalten als beispielsweise Baumarten gemäßigter Breiten.
Es zeigt sich, dass das Wasserleitsystem dieser Arten weniger kavitationsanfällig ist und dadurch höheren Wassermangel standhalten kann (Breda et al. 2006). Auch unsere Arten unterscheiden sich hinsichtlich der Kavitationsanfälligkeit, zum Beispiel liegt diese bei Fichte höher als bei Kiefer und Buche (Klein 2014). In der Schweiz konnte 2018 erstmalig ein solcher Kollaps der Wasserleitung messtechnisch »live« verfolgt werden. Während einige Fichten direkt abstarben, überlebten die Buchen, auch wenn sie einen Teil der Krone einbüßten (Schuldt et al. 2020). Ob »Verhungern « oder »Verdursten« den Ausschlag gibt, lässt sich dabei wohl nicht sagen. Beide Prozesse sind zu eng miteinander verknüpft (McDowell 2011).
Anpassungen an Trockenheit
- Schon die Anatomie der Blätter zeigt durch ihre wachsartige Cuticula, die Anordnung der Spaltöffnungen (Stomata) an der Blattunterseite (im Fall von Laubbäumen) und Verstärkung des Stützgewebes z.B. bei Sonnenblättern, wie wichtig ein Schutz vor unkontrollierter Verdunstung für den Baum ist.
- Durch Schließen der Stomata reduzieren Bäume kurzfristig und dynamisch ihren Wasserverlust. Allerdings verzichten sie damit auf Photosynthese, und auch die kühlende Wirkung der Verdunstung. Bäume nutzen oft den kühleren Vormittag, wo die Strahlung schon hoch, und die Luft noch nicht so trocken ist, für die Photosynthese und schließen mit zunehmendem Wasserverlust im Tagesverlauf die Stomata.
- Reduzierung der Blattfläche: einerseits durch Einrollen und/oder vorzeitiges Verfärben und Abwerfen (Seneszenz), andererseits durch Produktion von weniger und kleineren Blättern im Folgejahr. Im Gegensatz zum Stomataschluss ist dieser Prozess nicht reversibel, und die Erholung kann mehrere Jahre dauern.
- Verringerung der Verletzbarkeit der Leitungsbahnen für Lufteintritt (»Kavitation«) erfolgt langfristig u.a. mit kleineren Gefäßdurchmessern (Schuldt et al. 2016). Dieser Aspekt wird aktuell sehr intensiv erforscht. Neuere Ergebnisse deuten darauf hin, dass Laubbäume einen Verlust von bis zu 90 % ihrer Leitungsfläche überleben können, während Nadelbäume schon bei 50 % absterben (Urli et al. 2013).
- Durch die Fähigkeit, tiefer wurzeln zu können, erschließt sich ein Baum langfristig ein größeres Wasserreservoir im Boden und kann damit – sofern der Boden entsprechend Wasser speichert – Dürreperioden eine Zeit lang überbrücken. Auf trockenen Standorten und nach Dürrejahren investieren Bäume vermehrt in das Wurzelsystem. Das Wasser im Boden wird dabei nicht nur von den mikroskopisch kleinen Feinwurzeln aufgenommen, sondern auch über das weit verzweigte Geflecht der Mykorrhiza. Auch die Zusammensetzung der Mykorrhiza ändert sich infolge von Trockenheit.
- Erhöhung der Fähigkeit, dem Boden mehr Wasser entziehen zu können durch aktiv eingelagerte Stoffe im Blatt- und Wurzelgewebe; diese osmotischen Anpassung wurde u.a. bei den Gattungen Acer, Carpinus, Quercus, Prunus, Pinus und Picea gefunden (Ellenberg und Leuschner 2010).
Zusammenfassung
Diese Extreme sind die »Architekten« des Klimawandels, die sich in Arealverschiebungen von Baumarten und Arten allgemein manifestiert. Betroffen sind sowohl die Verjüngung als auch der Altbestand, die Auswirkungen sind standortsensitiv und wirken zusammen mit weiteren Faktoren.
Die Risiken unterscheiden sich je Baumart, da die Baumarten unterschiedlich stark an Wärme und Trockenheit angepasst sind. Da nicht vorhergesagt werden kann, wo und wann Extremjahre auftreten, ist man am besten gewappnet, wenn man seine Bestände schon früh mit wärme- und trockenheitstoleranten Arten anreichert und besonders gefährdete Standorte im Blick hat.
- Ackermann, J.; Adler, P.; Hoffman, K.; Hurling, R.; John, R.; Otto, L.-F.; Sagischewski, H.; Seitz, R.; Straub, C.; Stürtz, M. (2018): Früherkennung von Buchdruckerbefall durch Drohnen. AFZ-DerWald 19, S. 50–53
- BMEL - Bundesministerium für Ernährung und Landwirtschaft (Hrsg.) (2020): Ergebnisse der Waldzustandserhebung 2019. 60 S.
- Brandl, S.; Falk,W. (2019): Mortalität von Fichte und Buche – Einfluss von Klima und Mischung. AFZ-Der Wald 2, S. 10–13
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. (2006): Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 63(6), S. 625–644
- Ellenberg, H.; Leuschner, C. (2010): Vegetation Mitteleuropas mit den Alpen: in ökologischer, dynamischer und historischer Sicht. 6. Auflage. UTB, Stuttgart
- Hanewinkel, M.; Cullmann, D. A.; Schelhaas, M. J.; Nabuurs, G. J.; Zimmermann, N. E. (2013): Climate change may cause severe loss in the economic value of European forest land. Nature Climate Change, 3(3), 203-207
- Klein, T. (2014): The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Functional ecology, 28(6), S. 1313–1320
- Klemmt, H.-J.; Taeger, S.; Lemme, H.; Buras, A.; Straub, C.; Menzel, A. (2018): Absterbeerscheinungen der Kiefer in Mittelfranken. AFZ/ DerWald 11, S. 20–22
- Kölling, C.; Schmidt, O. (2013): Die Lüge der Überlebenden. LWF aktuell 96, S. 22–24 LWF - Bayerische Landesanstalt für Wald und Forstwirtschaft (2020): Wald unter Druck. LWF aktuell 125
- Hanewinkel, M.; Cullmann, D. A.; Schelhaas, M. J.; Nabuurs, G. J.; Zimmermann, N. E. (2013): Climate change may cause severe loss in the economic value of European forest land. Nature Climate Change, 3(3), S. 203–207
- Manion, P.D. (1981): Tree disease concepts. Prentice-Hall, Inc. Mauri, A.; Strona, G.; San-Miguel-Ayanz, J. (2017): EU-Forest, a high-resolution tree occurrence dataset for Europe. Scientific data, 4(1), S. 1–8
- McDowell, N.; Pockman, W. T.; Allen, C. D.; Breshears, D. D.; Cobb, N.; Kolb, T.; ... & Yepez, E. A. (2008): Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New phytologist, 178(4), S. 719–739
- McDowell, N. G. (2011): Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant physiology, 155(3), S. 1051–1059
- Raspe, S.; Schulz, C.; Kroll, F. (2003): Wenn schon im Sommer tonnenweise Blätter fallen. Lwf aktuell 43, S. 11–13
- Roloff, A. (2001): Baumkronen – Verständnis und praktische Bedeutung eines komplexen Naturphänomens. Ulmer, Stuttgart
- Schuldt, B., Knutzen, F., Delzon, S., Jansen, S., Müller-Haubold, H., Burlett, R., ... & Leuschner, C. (2016). How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction?. New Phytologist, 210(2), S. 443-458
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; ... & Kahmen, A. (2020): A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic and Applied Ecology
- StMELF - Bayerisches Staatsministerium für Ernährung, Landwirtschaft und Forsten (Hrsg.) (2019): Ergebnisse der Waldzustandserhebung 2019. 21 S.
- Thurm, E. A.; Hernandez, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Zlatanoc, T.M.; Hladnik, D.; Balic, B.; Freudenschuss, A.; Büchsenmeister, R.; Falk, W. (2018): Alternative tree species under climate warming in managed European forests. Forest Ecology and Management, 430, S. 485–497
- Urli, M.; Porté, A. J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. (2013): Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree physiology, 33(7), S. 672-683