Sie wächst und wächst und wächst - Fakten zum Wachstum der Buche - LWF Wissen 86
von Hans Pretzsch, Lehrstuhl für Waldwachstumskunde, Technische Universität München
Zusammenfassung: Langfristige Messungen zeigen ein hohes Zuwachsniveau der Buche (Fagus sylvatica L.), das insbesondere an den Rändern des Verbreitungsgebietes im Nordosten und im Gebirge weiter zunimmt. Auf warmen, trockenen Standorten, vor allem am Südwestrand des Verbreitungsgebiets, können die Zuwächse jedoch auch unter das bisherige Niveau absinken. Die Wachstumssteigerung mancherorts und die Wachstumsreduktion andernorts bilden keinen Widerspruch. Letztlich kommt es darauf an, ob sich die örtlichen Standortbedingungen zum ökologischen Optimum der Buche hin oder davon weg entwickeln und welche Anpassungsfähigkeit die Buche besitzt. Die Anpassungsfähigkeit der Buche an längerfristige Trockenheit, sowohl durch morphologische, physiologische und epigenetische Reaktionen auf Baumebene als auch durch Umverteilung von Zuwachs- und Mortalitätsmustern auf Bestandesebene, dürfte bisher deutlich unterschätzt worden sein. So konnten sich selbst 80-jährige Buchen an mehrjährige experimentelle Austrocknung anpassen und sich davon überraschend schnell wieder erholen. Angesichts der Klimaänderungen gilt es, die Resilienz und Resistenz von Beständen durch Strukturvielfalt und Mischung zu erhöhen. Das Potenzial der Buche für eine waldbauliche Anpassung an den Klimawandel wird nach hiesiger Übersicht unterbewertet und bisher nicht annähernd ausgeschöpft.
Einleitung
Das Wachstum der Buche (Fagus sylvatica L.) verändert sich im Bereich ihres ökologischen Optimums in Mitteleuropa (Pretzsch 2020, Spiecker et al. 2012) und insbesondere an den Verbreitungsgrenzen im nordöstlichen (Bolte et al. 2007) und südwestlichen Hügelland (Penuelas et al. 2007) und in den Gebirgen (Hilmers et al. 2019, Dulamsuren et al. 2017). Wir sehen mancherorts eine Steigerung (Bontemps und Esper 2011, Pretzsch et al. 2014) und in anderen Regionen einen Rückgang (Leuschner 2020) ihres Wachstums. Die zunächst widersprüchlich erscheinenden Reaktionsmuster werden verständlich, wenn die jeweiligen standörtlichen Ausgangsbedingungen, das Reaktions- und Anpassungsvermögen der Buche und die zurückliegende Bewirtschaftung in die Interpretation mit einbezogen werden.
Langfristige Versuchsflächen (Pretzsch et al. 2019) und Austrocknungsexperimente (Grams et al. 2021) geben eine Übersicht über das Wachstum der Buche in Abhängigkeit von Klimabedingungen und Trockenstressereignissen. Der Zuwachs von Bäumen und Beständen wird häufig als Indikator für Vitalität und Fitness verwendet (Dobbertin 2005). Allerdings haben Untersuchungen zum Stressverhalten von Bäumen in den letzten Jahren verdeutlicht, dass hohe Zuwächse nicht unbedingt auch hohe Abwehrkraft gegenüber abiotischen oder biotischen Stressfaktoren bedeuten (Matyssek et al. 2012). Vielmehr können hohe Zuwächse mit geringer Stresstoleranz und geringe Stammzuwächse mit hoher Stabilität und Persistenz einhergehen. Waren in der Vergangenheit in vielen Regionen hohe Zuwächse das Maß der Dinge, so ist man heute in manchen Regionen auch mit geringen Zuwächsen zufrieden, wenn nur der Wald überhaupt erhalten bleibt (Kölling et al. 2013). Diese veränderte Sichtweise wird auch bei der Diskussion des im Folgenden beschriebenen Wachstums der Buche berücksichtigt werden.
Langfristiger Wachstumstrend
Ohne ihre Reduktion durch den Menschen in der Vergangenheit (Mantel 1990) würde die Buche den Wald in Mitteleuropa dominieren. Gegenwärtig wird die Buche durch eine klimaangepasste Forstwirtschaft (Bowditch et al. 2020) mit Übergang zu naturnahen Mischwäldern wieder gefördert. Allerdings wird ihre Eignung für den Anbau unter in Zukunft trockeneren und wärmeren Bedingungen noch sehr unterschiedlich beurteilt (siehe u. a. Ammer et al. 2005, Bolte et al. 2010, 2007, Geßler et al. 2007, Rennenberg et al. 2004). Zur Orientierung wird hier zunächst der langfristige Wachstumsgang der Buche vorgestellt und das gegenwärtige mit ihrem historischen Wachstum verglichen.
Abbildung 1 zeigt die Entwicklung von Buchen (Stammdurchmesser und Baumbiomasse) in Abhängigkeit vom Alter und dem Kalenderjahr in den letzten Jahrhunderten. Die Auswertung gründet sich auf 591 Buchen mit einem Alter bis zu 469 Jahren, Durchmessern bis zu 81,4 cm und Baumbiomassen bis 6,7 t. Die Zuwachsanalysen reichen bis in das Jahr 1546 zurück. Alle Bäume wurden in Deutschland beprobt (Pretzsch 2020) und schließen sowohl arme, trockene Standorte als auch nährstoffreiche, feuchte Standorte mit Bonitäten von 22 – 40 m im Alter 100 (Schober 1967, mä. Df.) ein. Aus der Abbildung geht hervor, dass die Steigung der Wachstumskurven von 1850 bis 2000 deutlich zugenommen hat. Beispielsweise erreichten Buchen im Alter von 50 Jahren im Jahr 1900 einen durchschnittlichen Durchmesser von 14,2 cm, im Jahr 1950 einen Durchmesser von 18,9 und in 2000 einen Durchmesser von 24,9 cm. Die analogen Massenwerte sind 0,104, 0,206, und 0,401 Tonnen in den Jahren 1900, 1950 bzw. 2000. Die Beschleunigung des Wachstums hat in den letzten Jahrzehnten leicht abgenommen. Auf das Durchmesserwachstum bezogen, betrug sie 34 % von 1850 – 1900 und 32 % in den letzten 50 Jahren. Ebenso war die Steigerung des Biomassenwachstums vor 200 – 300 Jahren am größten; gegenwärtig ist sie immer noch hoch, aber tendenziell abnehmend. Diese leichte Abnahme der Wachstumssteigerung (also nicht etwa Abnahme des Wachstums) gegenüber historischen Bedingungen dürfte auf eine Annäherung der Buche an ihre optimalen Wuchsbedingungen zurückzuführen sein (Pretzsch et al. 2014, Spiecker 2000). Bosela et al. (2016), Dulamsuren et al. (2017) und Penuelas et al. (2007) diskutieren eine mögliche Überschreitung des Optimums und Abnahme des Wachstums bei zunehmender Erwärmung und Trockenheit in Mittel- und Südeuropa.
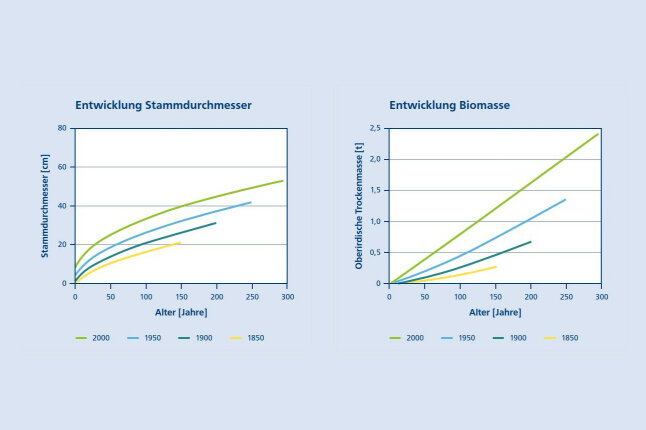
Abbildung 1a (links) und 1b (rechts): Entwicklung von (1a) Stammdurchmesser und (1b) Biomasse von Buchen in Abhängigkeit vom Baumalter und Kalenderjahr. Das Baumalter wurde durch Jahrringzählungen in der Baumhöhe von 1,30 m bestimmt. Die Kurvenverläufe zeigen den Stammdurchmesser bzw. die Baummasse, die die Buchen in gleichem Alter in den Jahren 1850, 1900, 1950 und 2000 erreichten (© LWF)
Dieser auf Baumebene gefundene Wachstumstrend zeigt sich auch auf der Bestandesebene (Abb. 2). Die Entwicklung der Gesamtwuchsleistung und des durchschnittlichen Gesamtzuwachses, dGZ, an Derbholzvolumen weicht auf vielen der in Abbildung 2 zusammengestellten 97 langfristigen Versuchsflächen in Europa deutlich positiv von den Erwartungswerten der Ertragstafel ab. Bei den langfristigen Entwicklungen liegen mehr als die Hälfte der dargestellten A-Grad Flächen sogar über der I. Bonität der Ertragstafel. Auf manchen Flächen wird das Niveau der Ertragstafeln aber auch deutlich unterschritten. Zahlreiche Untersuchungen sehen die Erhöhung der Jahrestemperatur, die Verlängerung der Vegetationszeit und die vermehrten Stickstoffeinträge als wesentliche Ursachen für die Wuchsbeschleunigung (Pretzsch et al. 2014, Tegel et al. 2014, Spiecker et al. 2012). Diese führen insbesondere in ausreichend wasserversorgten mittel- und nordeuropäischen Regionen, wo das Wachstums bisher durch Klima oder Nährstoffversorgung limitiert war, zu Zuwachsanstiegen. In mittel- und südeuropäischen Regionen mit zunehmender Sonneneinstrahlung, Wasserlimitierung, oder Spätfrosthäufigkeit kann das Wachstum der Buche insbesondere an der südwestlichen Verbreitungsgrenze auch deutlich rückläufig sein (Penuelas et al. 2007). Besonders deutlich sind die Wachstumssteigerungen in jüngeren Beständen, also im linken oberen Bereich der Kurvenbündel in Abbildung 2, ausgeprägt. Gleichzeitig können auf besonders trockenen Standorten aber auch Zuwachseinbußen und eine vermehrte Mortalität beobachtet werden (Leuschner 2020). Inwieweit die vermehrten Dürreschäden in Buchenbeständen auch mit der waldbaulichen Behandlung zusammenhängen, beispielsweise mit der abrupten Auflichtung in älteren Beständen, ist Gegenstand neuerer und laufender Untersuchungen (Meyer et al. 2022, Pretzsch 2021). Wachstumssteigerung und Wachstumsreduktion bilden also keinen Widerspruch, letztlich kommt es darauf an, ob sich die Standorte zum ökologischen Optimum der Buche hin oder davon weg entwickeln.
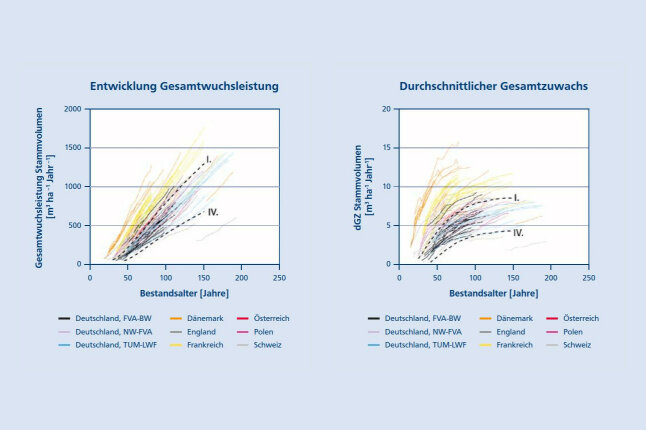
Abbildung 2a (links) und 2b (rechts): Entwicklung der Gesamtwuchsleitung (2a) und des durchschnittlichen Gesamtzuwachses, dGZ an Derbholzvolumen (2b) auf 97 langfristigen Versuchsflächen in verschiedenen europäischen Ländern nach Pretzsch et al. (2019). Die Kurvenverläufe reichen bis 1871 zurück und sind im Vergleich zur Ertragstafel von Schober (1967) Bonität I. und IV für mäßige Durchforstung dargestellt (© LWF)
Veränderung im Wuchsverhalten der Buche zeigt sich insbesondere im Gebirge. Basierend auf Bohrkernen von 660 Buchen auf 28 langfristigen Versuchsflächen in Bergwäldern sieben europäischer Länder, von Bulgarien im Südosten bis zur Schweiz und Deutschland im Norden, konnten Pretzsch et al. (2020) eine charakteristische Veränderung des Durchmesserwachstums der Buche in unterschiedlichen Höhenlagen in den letzten 300 Jahren zeigen. Die Untersuchungsbestände lagen zwischen 621 – 1.569 m über NN, mit Jahresmitteltemperaturen von 2,9 °C – 8,2 °C und Jahresniederschlägen von 794 – 2.767 mm. Das untere, mittlere und obere Kurvenbündel in Abbildung 3a zeigt den Entwicklungsgang von Buchen, die im Kalenderjahr 1700, 1800 bzw. 1900 eine Höhe von 1.30 m hatten (hier genannt dKalenderjahr). Demnach haben sich die Wuchsbedingungen in den letzten Jahrhunderten derart verbessert, dass die Buchen in Höhenlagen von 1.000 – 1.300 m über NN gegenwärtig besser wachsen als in 700 m Höhe, während das vor 200 – 300 Jahren noch andersherum war.
Abbildung 3b zeigt das Wachstum der Buche im Gebirge in Reinbeständen (durchgezogene Linien) im Vergleich zu Mischbeständen (gestrichelt) in Höhenlagen von 700 – 1.300 über NN. Die Untersuchung basiert auf Bohrkernuntersuchungen an 1.240 dominanten Buchen auf Versuchsflächen in 45 Rein- und 46 Mischbeständen in Bergwäldern, in 14 europäischen Ländern von der Schweiz im Westen über u. a. Deutschland, Italien, Tschechien, Polen, die Ukraine und Rumänien bis Bulgarien im Südosten. Die Abbildung zeigt sowohl in Reinbeständen (durchgezogene Linien) als auch in Mischbeständen (gestrichelte Linien) eine Abnahme des Wachstums der Buche von der Höhenlage 700 bis 1.300 m über NN (Pretzsch et al. 2021). Diese Abnahme ist allerdings in Mischbeständen wesentlich geringer ausgeprägt, und in einer Höhenlage oberhalb 1.200 m über NN ist das Wachstum der Buche in Mischbeständen sogar höher als in Reinbeständen. Offenbar fördert in dieser Höhenlage die Mischung, z. B. durch Reduktion von Frostschäden, das Wachstum der Buche.
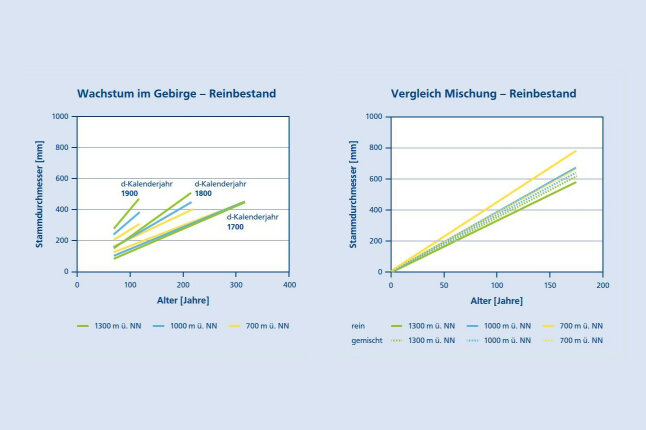
Abbildung 3a (links) und 3b (rechts): Wachstum der Buche im Gebirge in Reinbeständen (3a) und in Mischung im Vergleich zu Reinbeständen (3b). (3a) Das untere, mittlere und obere Kurvenbündel zeigt den Entwicklungsgang von Buchen, die im Kalenderjahr 1700, 1800 bzw. 1900 eine Höhe von 1.30 m hatten (Pretzsch et al. 2020). (3b): Wachstumsgang der Buche in Reinbeständen (durchgezogene Linien) im Vergleich zu Mischbeständen (gestrichelt) in verschiedenen Höhenlagen europäischer Gebirge (Pretzsch et al. 2021) (© LWF)
Stressreaktionen auf hohem Niveau
Aus zahlreichen Untersuchungen ist bekannt, dass einzelne Trockenjahre, wie z. B. 1976, 2003, oder 2015 den Zuwachs von Buchen deutlich weniger reduzieren als den Zuwachs von Fichten (Picea abies [L.] Karst.; Pretzsch et al. 2013, Zang et al. 2011 a und b). Im Vergleich zur Eiche (Quercus petraea (MATT.)) schneidet die Buche aber schlechter ab (Abb. 4a). In Reinbeständen hat die Buche im Vergleich zu anderen Baumarten eine mittlere Resistenz aber auch eine mittlere Erholungsfähigkeit. In Mischung mit Buche reagieren Fichte und Eiche auf Trockenheit ähnlich wie in Reinbeständen. Die Buche hingegen ist in diesen Mischbeständen signifikant resistenter und resilienter als im Reinbestand (Abb. 4b). Insbesondere in Mischung mit Eiche kann sich der Trockenstress der Buche verringern, ohne dass die Eiche dadurch zusätzliche Zuwachseinbußen erleidet (Abb. 4c).
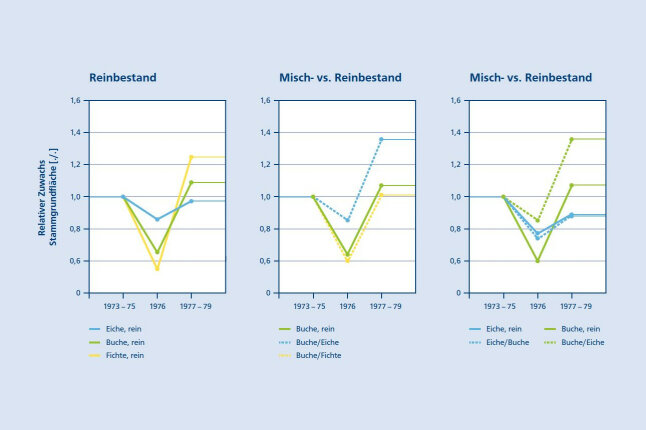
Abbildung 4a (links), 4b (Mitte) und 4c (rechts): Artspezifische Zuwachsreaktionen von Buche, Fichte und Eiche im Trockenjahr 1976 in Relation zum mittleren Zuwachs in der vorherigen dreijährigen Referenzperiode 1973 – 1975 (nach Pretzsch et al. 2013). (4a): Reaktionsmuster von Buche, Fichte und Eiche in Reinbeständen, (4b): Verhalten der Buche in Misch- im Vergleich zu Reinbeständen, (4c): Buche und Eiche in Misch- im Vergleich zu Reinbeständen. Die Kurvenverläufe zeigen den Zuwachs im Trockenjahr 1976 und in der Erholungsphase 1977 – 1979 in Relation zum mittleren Zuwachs in der Referenzperiode von 1973 – 1975 (© LWF)
Dendrochronologische Untersuchungen konzentrieren sich häufig auf die relativen Schwankungen des Zuwachsganges und eliminieren, d. h. vernachlässigen, das absolute Zuwachsniveau. Deshalb zeigt Abbildung 5 sowohl das absolute Zuwachsniveau in den zurückliegenden 20 Jahren, als auch die Zuwachsreaktionen auf die Trockenjahre 2003 und 2015. Ausgewertet wurden hier die Zuwachsverläufe von 54 mittelalten, vollbestockten und nicht oder nur schwach durchforsteten Reinbeständen aus Buchen, Kiefern (Pinus sylvestris L.), Fichten und Eichen (aus Platzgründen nicht gezeigt). Die gemessenen jährlichen Volumenzuwächse liegen um 5 – 77 % und im Mittel um 48 % über den jeweiligen Ertragstafeln. Besonders deutlich sind die positiven Abweichungen von den Ertragstafeln bei Eiche, Kiefer und Buche (+ 47 – 77 %) ausgeprägt. In den zuletzt genannten Beständen kommt es, im Vergleich zum langfristigen Trend, in Jahren mit großer Trockenheit zwar zu Einbrüchen um 19 – 22 %, die erreichten Zuwächse liegen aber immer noch über den von den Ertragstafeln prognostizierten Werten. Das bedeutet, dass die Zuwächse in Extremjahren zwar deutlich unter das vorherige Niveau absinken, dass sie aber trotz dieser Verluste im Durchschnitt noch immer über den historischen Werten der Ertragstafeln verlaufen. Nur in wenigen ungünstigsten Fällen sinken die Zuwächse in Trockenjahren unter das Niveau der Ertragstafeln ab. In Buchenbeständen liegen die Zuwächse auch in Trockenjahren durchschnittlich noch immer bei 133 % der Ertragstafel. D. h. sie sinken im Vergleich zum langfristigen Trend ab, liegen aber noch immer deutlich über den langfristigen Werten der Ertragstafeln (Abb. 5a).
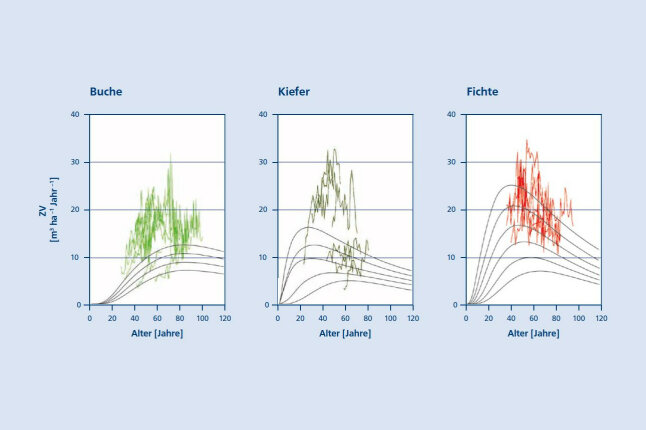
Abbildung 5a (links), 5b (Mitte) und 5c (rechts): Verlauf des absoluten jährlichen Volumenzuwachses (Vorratsfestmeter Derbholz mit Rinde m3 ha-1 Jahr-1) in Reinbeständen aus (5a) Buche, (5b) Kiefer und (5c) Fichte auf ungünstigen bis günstigen Standorten in Deutschland. Dargestellt sind die Zuwachsverläufe im Zeitraum 1997 – 2018 im Vergleich zu den Ertragstafeln von Schober (1967), Wiedemann (1943) und Assmann and Franz (1963) (nach Pretzsch et al. 2020)
Anpassung auf Baum- und Bestandesebene
Besonders wenig bekannt ist bisher über die Anpassungsfähigkeit der Buche an mehrjährigen Trockenstress, wie er unter zukünftigen Klimabedingungen zu erwarten ist. Interessante Einblicke in die zu erwartenden Reaktionsmuster bietet hier das vor zehn Jahren angelegte Dachexperiment KROOF im Kranzberger Forst bei Freising (Grams et al. 2021). Dort wurden sechs Parzellen mit etwa 80jährigen Fichten und Buchen im Rein- und Mischbestand durch Überdachung ausgetrocknet (Regenausschluss A) und mit sechs benachbarten Parzellen (Kontrolle K) verglichen. Die Austrocknung erfolgte von 2014 bis 2018, also über fünf Jahre hinweg. Die Dächer mit regengesteuertem Schließmechanismus waren während der Vegetationsperiode von April bis Oktober bei Niederschlagsereignissen geschlossen. Über Winter wurde der Niederschlag zur Auffüllung des Bodenwasserspeichers zugelassen. Seit 2016 werden die Fichten, sowohl auf den Kontroll- als auch auf den Regenausschlussflächen, jährlich im Frühjahr, zur Vermeidung von Borkenkäferbefall, mit einem Insektizid besprüht, nachdem einige befallene Fichten entfernt werden mussten. Im Sommer 2019 wurden die Parzellen geringfügig bewässert.
Der Effekt der Austrocknung im Jahr 2014 zeigt sich durch eine deutliche Abnahme des Radialzuwachses bei den ausgetrockneten Fichten und Buchen (Pretzsch et al. 2014, 2020). Der Radialzuwachs der ausgetrockneten Buche ging weniger stark zurück als jener der Fichte und pendelte sich schneller unter dem Niveau der Kontrolle ein (Abb. 6a). Eine deutlich erkennbare Anpassung und Erholung setzte schon vor Beginn der Bewässerung 2019 ein, und der Radialzuwachs der behandelten Bäume erreichte 2019 schon wieder fast das Niveau der Kontrollgruppe. Im Mittel fiel der Radialzuwachs der Fichte mit Beginn der Austrocknung steil unter das Niveau der Kontrolle, stabilisierte sich dann auf geringem Niveau, bis zu einer erkennbaren Erholung mit Beginn der Bewässerung im Jahr 2019 (Abb. 6b). Während der Austrocknung (2014 – 2018) erreichte die Fichte nur etwa die Hälfte des Radialzuwachses der Kontrollgruppe und blieb auch nach der Bewässerung deutlich hinter den Kontrollbäumen zurück. Das KROOF-Experiment zeigt, wie der Zuwachs von Fichten und Buchen auf längerfristige Trockenheit reagiert, wie die Artenmischung die Zuwachsreaktionen modifiziert, und wie sich beide Arten bei anschließender Bewässerung erholen.
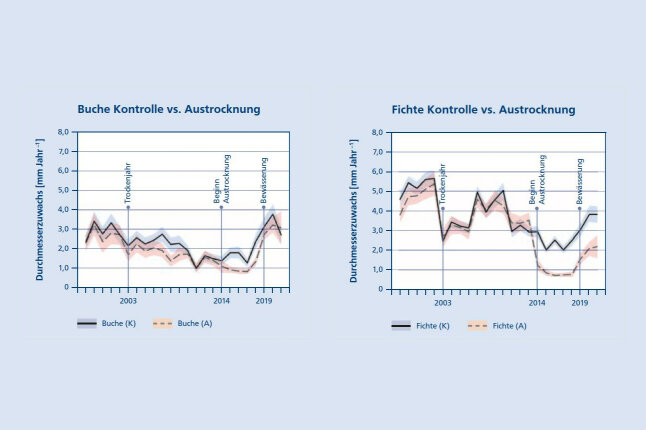
Abbildung 6a (links) und 6b (rechts): Mittlerer jährlicher Durchmesserzuwachs (± Standardfehler) der Einzelbäume der Buche (6a) und der Fichte (6b) auf den ausgetrockneten Parzellen (A) im Vergleich zu den Kontrollflächen (K). Das Trockenjahr 2003, der Beginn der Austrocknung 2014, sowie das Ende der Austrocknung mit Bewässerung im Jahr 2019 sind durch vertikale Linien hervorgehoben (© LWF)
Abbildung 7a zeigt, dass der Volumenzuwachs des Buchenreinbestandes auf dem KROOF Experiment durch die Austrocknung von 7,0 auf im Mittel 4,0 m³ ha⁻¹ Jahr⁻¹, also um 43 % zurückgegangen ist. Der in den Jahren 2014 – 2018 akkumulierte Zuwachsverlust beträgt bei der Buche 15,2 m ha⁻¹. In dem Fichtenreinbestand (Abb. 7b) hat der Volumenzuwachs durch die Austrocknung im Mittel von 9,6 auf 3,6 m³ ha⁻¹ Jahr⁻¹, also um 63 % abgenommen. In der fünfjährigen Trockenperiode akkumulierte sich ein Zuwachsverlust von 30,4 m³ ha⁻¹. Das Ende der Austrocknung mit Bewässerung im Jahr 2019 äußerte sich bei beiden Baumarten in einer deutlichen Aufwärtsbewegung des Zuwachsganges, wobei der Fichtenbestand das Niveau der Kontrollfläche bei weitem nicht wieder erreichte. Der Zuwachs des zuvor ausgetrockneten Buchenbestandes stieg wieder annähernd auf das Niveau der Kontrollfläche an. Die Untersuchung unterstreicht, dass der Zuwachsrückgang in einzelnen Trockenjahren und im ersten Jahr der Austrocknung besonders deutlich ausfällt, dann bei längerer Austrocknung aber eine gewisse Stabilität erkennen lässt. Selbst nach fünfjähriger Austrocknung zeigen beide Baumarten, insbesondere die Buche, die Fähigkeit zur Erholung und eine Annäherung an das alterstypische Zuwachsniveau (vgl. rot eingetragene Linien der Ertragstafeln von Schober (1967) bzw. Assmann und Franz (1963)).
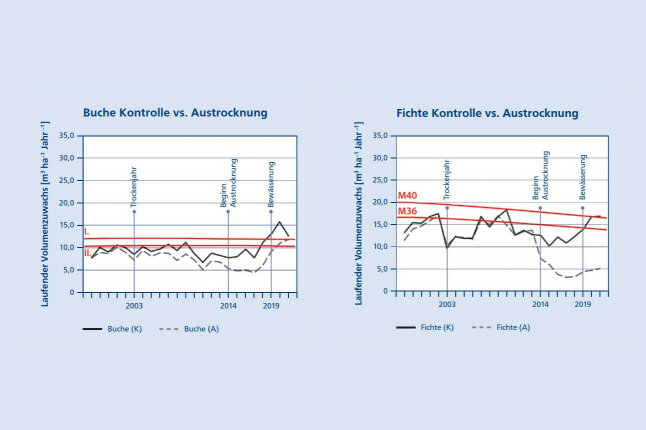
Abbildung 7a (links) und 7b (rechts): Laufender jährlicher Volumenzuwachs pro ha der (7a) Buche im Vergleich zur Fichte (7b) auf den ausgetrockneten Parzellen (A) im Vergleich zu den Kontrollflächen (K). Zusätzlich ist in Rot das Zuwachsniveau der Ertragstafel (ET für Buche I. und II. nach Schober (1975); ET für Fichte M40 und M36 nach Assmann und Franz (1963)) dargestellt. Das Trockenjahr 2003, der Beginn der Austrocknung 2014, sowie das Ende der Austrocknung mit Bewässerung im Jahr 2019 sind durch vertikale Linien hervorgehoben (© LWF)
Auch auf Bestandesebene können Buchen Zuwachseinbußen in Trockenjahren in gewissem Maße abpuffern. Herrschende Buchen haben eine deutlich geringere Trockenresistenz als zwischen- und unterständige Bäume (Abb. 8a). Dieses günstigere Abschneiden kleinerer Bäume ist in Fichtenbeständen weniger deutlich als in den Buchenbeständen (Abb. 8b). Eine Position in der Zwischen- und Unterschicht bei Trockenheit ist für Fichten demnach weniger vorteilhaft als für Buchen. Bei der Buche liegen der in Abbildung 8 dargestellten Untersuchung die Bestände der Waldklimamessstationen in Bayern Bad Brückenau, Ebrach, Mitterfels und Schongau zugrunde (Intensive Forest Monitoring, Level II, vgl. Ferretti und Fischer 2013). Diese Bestände sind mit jährlich abgelesenen Dauerumfangmessbändern ausgestattet und ermöglichen deshalb die Berechnung des Grundflächenzuwachses aller Bäume im Trockenjahr 2003 und auch ihres mittleren jährlichen Zuwachses in den vorhergehenden normalen Jahren 2000 – 2002. Der Quotient zwischen dem Zuwachs im Trockenjahr und der Referenzperiode 2000 – 2002 erbringt die Resistenz, die in Abbildung 8 auf etwaige Abhängigkeit von der Ausgangsgrundfläche zu Periodenbeginn (Ausgleichslinien in Abb. 8) analysiert wird.
Wäre der Zuwachs von Bäumen aller Größen im Trockenjahr nicht reduziert, so würden die Ausgleichslinien auf dem Niveau 1.0 und damit parallel zur x-Achse verlaufen. Wäre der Zuwachs aller Bäume gleichermaßen durch die Trockenheit reduziert, so verliefen die Linien ebenfalls parallel zur x-Achse, aber auf einem reduzierten Niveau, also unterhalb der 1.0-Linie. Die Untersuchung dieser Zusammenhänge ergab, dass die Resistenz bei kleinen Bäumen am höchsten ist und insbesondere in Buchenbeständen mit der Größe abnimmt (Abb. 8a). Offenbar haben subdominante Buchen geringeren hydraulischen Stress, ziehen Vorteile aus ihrer Beschattung und profitieren von dem eingeschränkten Wasserverbrauch ihrer größeren und deshalb sonnenexponierten Nachbarn. Auf diese Weise können kleine Bäume den Zuwachsverlust ihrer größeren Nachbarn teilweise kompensieren und den Bestandeszuwachs in Trockenjahren stabilisieren. Diese stabilisierende Rolle kleinerer Bäume wird bei den zumeist auf dominante Bäume beschränkten Stressuntersuchungen übersehen. Andererseits können kleine Bäume in Beständen insbesondere aufgrund ihrer meist großen Anzahl auch die Wasserversorgung großer Bäume reduzieren und deren Trockenstress weiter verstärken. Ob die Entnahme kleiner Bäume sich auf den Bestandeszuwachs letztlich positiv (wegen Entlastung des Wasserverbrauchs) oder negativ (wegen Wegfall ihrer Zuwachsstabilisierung aufgrund höherer Resistenz) auswirkt, ist Gegenstand laufender Untersuchungen.
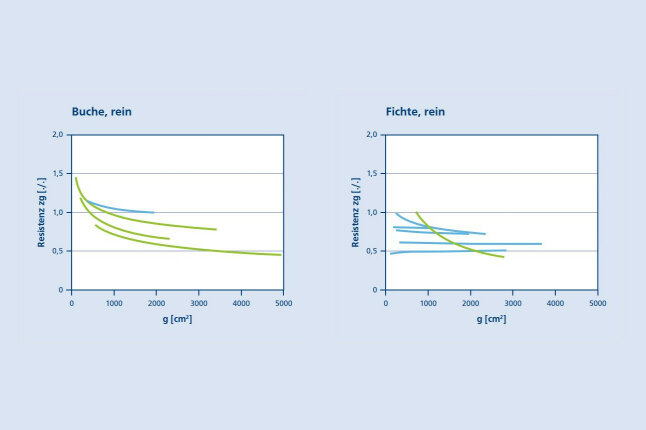
Abbildung 8a (links) und 8b (rechts): Zusammenhang zwischen der Stammgrundfläche zu Periodenbeginn und der Trockenresistenz im Jahr 2003 auf den Waldklimamessstationen in Bayern in Reinbeständen aus Buche (8a) und Fichte (8b). Kurven mit einer signifikanten Abnahme der Trockenresistenz mit zunehmender Baumgröße sind als grüne Linien eingetragen, nicht signifikant ab- oder zunehmende Kurvenverläufe sind blau hervorgehoben (Pretzsch et al. 2022)
Perspektiven
Ein »Abschreiben« der Buche angesichts des Klimawandels zugunsten fremdländischer Baumarten, deren Möglichkeiten und Grenzen im Anbau noch nicht wissenschaftlich abgesichert sind, erscheint angesichts des hier dargestellten Zuwachsverhaltens der Buche als falsch. In trockenen Lagen in ihrem Verbreitungsgebiet und an dessen Rand ermöglichen Mischbestände eine Risikostreuung und Stabilisierung der Buche aufgrund ihrer Förderung durch benachbarte Baumarten wie beispielsweise der Eiche. In Mischbeständen mit Koniferen oder Eiche kann die Buche wirkungsvoll zu Erhöhung und Stabilisierung des Zuwachses beitragen (Pretzsch et al. 2013, Pretzsch und Schütze 2021). Die Beimischung von Buche kann dort auch zur wirksamen Reduktion von Sturmwürfen, Spätfrostschäden oder biotischen Kalamitäten beitragen. Die bisherigen Untersuchungen zum Stressverhalten bei Trockenheit beziehen sich meist auf einzelne Extremjahre. Untersuchungen zu Anpassung an trockenere Wuchsbedingungen stehen noch ganz am Anfang. Erste Ergebnisse deuten aber darauf hin, dass die insgesamt sehr plastische Buche auch in ihrer physiologischen, morphologischen und epigenetischen Anpassungsfähigkeit unterschätzt wird. Auch eine kontinuierlichere waldbauliche Behandlung mit einer Vermeidung wiederholter starker und abrupter Freistellung könnte die Vitalität erhöhen. Auf trockenen Standorten können zwischen- und unterständige Bäume Zuwachseinbußen, die vor allem herrschende Buchen in Trockenjahren erleiden, teilweise abpuffern. Strukturreiche Buchenbestände, Buchenplenterwälder oder Mischbestände mit klimatoleranteren Koniferen wie Kiefer, Tanne oder Douglasie haben nach hiesigen Ergebnissen ein hohes Zukunftspotenzial. Über die Klimastabilität hinaus erbringen solche Bestände die vielfältigen ökonomischen, ökologischen und sozioökonomischen Funktionen und Leistungen, wie sie für die integrative Forstwirtschaft in Mitteleuropa essenziell sind.
Danksagung
Dank geht an das Bayerische Staatsministerium für Ernährung, Landwirtschaft und Forsten für die langjährige Förderung des Ertragskundlichen Versuchswesens in Bayern (Projekt W 07) und an die Bayerischen Staatsforsten (BaySF) für die Unterstützung der Versuchsflächenarbeit. Gedankt wird auch Frau Bradatsch für die Anfertigung der Grafiken und Frau Felsl für das Korrekturlesen und Formatieren.
Literatur
- Ammer, C.; Albrecht, H.; Borchert, H.; Brosinger, F.; Dittmar, C.; Elling, W.; Ewald, J.; Felbermeier, B.; von Gilsa, H.; Kenk, G., et al. (2005): Zur Zukunft der Buche (Fagus sylvatica L.) in Mitteleuropa. Kritische Anmerkungen zu einem Beitrag von Rennenberg et al. Allg. Forst-u. J.-Ztg., 176(4): S–60.
- Assmann, E.; Franz, F.; (1963): Vorläufige Fichten-Ertragstafel für Bayern. Forstl Forschungsanst München, Inst Ertragskd, 104 S.
- Bolte, A.; Czajkowski, T.; Kompa, T. (2007): The north-eastern distribution range of European beech—a review. Forestry, 80(4), 413-429.
- Bolte, A.; Hilbrig, L.; Grundmann, B.; Kampf, F.; Brunet, J.; Roloff, A. (2010): Climate change impacts on stand structure and competitive interactions in a southern Swedish spruce–beech forest. European Journal of Forest Research, 129(3), 261-276.
- Bosela, M.; Štefančík, I.; Petráš, R.;Vacek, S. (2016): The effects of climate warming on the growth of European beech forests depend critically on thinning strategy and site productivity. Agricultural and Forest Meteorology, 222, 21-31.
- Bowditch, E.; Santopuoli, G.; Binder, F.; del Rio, M.; La Porta, N.; Kluvankova, T.; Lesinski, J.; Motta, R.; Pach, M.; Panzacchi, P.; Pretzsch, H.; Temperli, Ch.; Tonon, G.; Smith, M.; Velikova, V.; Weatherall, A.; Tognetti, R.; (2020): What is Climate Smart Forestry? A definition from a multinational collaborative process focused on mountain regions of Europe. Ecosystem Services. 43, 101113. https://doi.org/10.10/j.ecoser.2020.101113.
- Dobbertin, M. (2005): Tree growth as indicator of tree vitality and of tree reaction to environmental stress: a review. European Journal of Forest Research, 124(4), 319-333.
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. (2017): European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees, 31(2), 673-686.
- Ferretti, M.; Fischer, R. (2013): Forest Monitoring: Methods for terrestrial investigations in Europe with an overview of North America and Asia. Developments in Environmental Science, Vol. 12. Elsevier, Oxford, 507 p.
- Geßler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. 2007: Potential risks for European beech (Fagus sylvatica L.) in a changing climate. Trees 21(1): 1–11. doi:10.1007/s00468-006-0107-x.
- Grams, T. E.; Hesse, B. D.; Gebhardt, T.; Weikl, F.; Rötzer, T.; Kovacs, B.; ... Pritsch, K. (2021): The Kroof experiment: realization and efficacy of a recurrent drought experiment plus recovery in a beech/spruce forest. Ecosphere, 12(3), e03399.
- Hilmers, T.; Avdagić, A.; Bartkowicz, L.; Bielak, K.; Binder, F.; Bončina, A.;... Pretzsch, H. (2019): The productivity of mixed mountain forests comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. Forestry: An International Journal of Forest Research, 92(5), 512-522.
- Kölling, C.; Binder, F.; Falk, W. (2013): Risiko und Ertrag in ungewisser Zukunft: Der Klimawandel fordert die Generationengerechtigkeit heraus. Lwf Wissen, 72, 54-58.
- Leuschner, C. (2020): Drought response of European beech (Fagus sylvatica L.) - A review. Perspectives in Plant Ecology, Evolution and Systematics, 47, 125576.
- Mantel, K. (1990): Wald und Forst in der Geschichte: ein Lehr-und Handbuch. Schaper, Hannover.
- Matyssek, R.; Schnyder, H.; Oßwald, W.; Ernst, D.; Munch, J. C.; Pretzsch, H. (2012): Growth and defence in plants. Ecological Studies, 220.
- Meyer, P.; Spînu, A. P.; Mölder, A.; Bauhus, J. (2022): Management alters drought‐induced mortality patterns in European beech (Fagus sylvatica L.) forests. Plant Biology. doi:10.1111/plb.13396
- Penuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. (2007): Migration, invasion and decline: changes in recruitment and forest structure in a warming‐linked shift of European beech forest in Catalonia (NE Spain). Ecography, 30(6), 829-837.
- Pretzsch, H. (2020): The course of tree growth. Theory and reality. Forest Ecology and Management, 478, 118508.
- Pretzsch, H. (2021): Trees grow modulated by the ecological memory of their past growth. Consequences for monitoring, modelling, and silvicultural treatment. Forest Ecology and Management, 487, 118982. https://doi.org/10.1016/j.foreco.2021.118982
- Pretzsch, H.; Schütze, G. (2021): Tree species mixing can increase stand productivity, density, and growth efficiency and attenuate the tradeoff between density and growth throughout the whole rotation. Annals of Botany, Annals of Botany, Volume 128, Issue 6, 2 November 2021, Pages 767–786
- Pretzsch, H.; Ammer, C.; Wolff, B.; Steckel, M.; Rukh, S.; Heym, M. (2020): Zuwachsniveau, Zuwachstrend und episodische Zuwachseinbrüche. Ein zusammenfassendes Bild vom aktuellen Zuwachsgang in Rein-und Mischbeständen aus Fichte, Kiefer, Buche und Eiche. Allgemeine Forst-u. Jagdzeitung, 191(1/2), 21.
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. (2014): Forest stand growth dynamics in Central Europe have accelerated since 1870. Nature communications, 5(1), 1-10.
- Pretzsch, H.; del Río, M.; Biber, P.; Arcangeli, C.; Bielak, K.; Brang, P.; ... Sycheva, E. (2019): Maintenance of long-term experiments for unique insights into forest growth dynamics and trends: review and perspectives. European Journal of Forest Research, 138(1), 165-185.
- Pretzsch, H.; del Río, M.; Grote, R.; Klemmt, H.-J.; Ordóñez, C.; Bravo Oviedo F. (2022): Tracing drought effects from the tree to the stand growth in temperate and Mediterranean forests. Insights and consequences for forest ecology and management, Trees. Structure and Function, submitted.
- Pretzsch, H.; Hilmers, T.; Biber, P.; Avdagić, A.; Binder, F.; Bončina, A.; ... Tognetti, R. (2020): Evidence of elevation-specific growth changes of spruce, fir, and beech in European mixed mountain forests during the last three centuries. Canadian Journal of Forest Research, 50(7), 689-703.
- Pretzsch, H.; Hilmers, T.; Uhl, E.; Bielak, K.; Bosela, M.; del Rio, M.; ... Tognetti, R. (2021): European beech stem diameter grows better in mixed than in mono-specific stands at the edge of its distribution in mountain forests. European Journal of Forest Research, 140(1), 127-145.
- Pretzsch, H.; Motte, F.; Grams, Th.; Prtisch, K.; Rötzer, Th.; Hesse, B.; Häberle, K-H. (2021): Fichten und Buchen bei Trockenheit im Rein- und Mischbestand, AFZ Der Wald, 20: 12-15
- Pretzsch, H.; Schütze, G.; Uhl, E. (2013): Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter‐specific facilitation. Plant Biology, 15(3), 483-495.
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H. P.; ... Zingg, A. (2013): Productivity of mixed versus pure stands of oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.) and European beech (Fagus sylvatica L.) along an ecological gradient. European Journal of Forest Research, 132(2), 263-280.
- Rennenberg, H.; Seiler, W.; Matyssek, R.; Gessler, A.; Kreuzwieser, J. (2004): Die Buche (Fagus sylvatica L.) - ein Waldbaum ohne Zukunft im südlichen Mitteleuropa. Allgemeine Forst-und Jagdzeitung 175(10–11): 210–224.
- Schober, R. (1967): Buchen-Ertragstafel für mäßige und starke Durchforstung, In: Schober R (1972) Die Rotbuche 1971. Schr Forstl Fak Univ Göttingen u Niedersächs Forstl Versuchsanst 43/44, JD Sauerländer’s Verlag, Frankfurt am Main, 333 S.
- Schober, R. (1975): Ertragstafeln wichtiger Baumarten. JD Sauerländer’s Verlag, Frankfurt am Main
- Spiecker, H.; Mielikäinen, K.; Köhl, M.; Skovsgaard, J. P. (Eds.). (2012): Growth trends in European forests: studies from 12 countries. Springer Science & Business Media.
- Spiecker, H. (2000): Growth of Norway spruce (Picea abies [L.] Karst.) under changing environmental conditions in Europe. In Spruce Monocultures in Central Europe. Problems and Prospects”. EFI Proceedings (No. 33, pp. 11-26).
- Tegel, W.; Seim, A.; Hakelberg, D.; Hoffmann, S.; Panev, M.; Westphal, T.; Büntgen, U. (2014): A recent growth increase of European beech (Fagus sylvatica L.) at its Mediterranean distribution limit contradicts drought stress. European Journal of Forest Research, 133(1), 61-71.
- Wiedemann, E. (1943): Kiefern-Ertragstafel für mäßige Durchforstung, starke Durchforstung und Lichtung, In: Wiedemann E (1948) Die Kiefer 1948. Verlag M & H Schaper, Hannover, 337 S.
- Zang, C.; Rothe, A.; Weis, W.; Pretzsch, H. (2011a): Tree suitability under climate change conditions: susceptibility of major forest tree species from tree-rings widths. Allg Forst u J Ztg, 182, 98-112.
- Zang, C.; Rothe, A.; Weis, W.; Pretzsch, H. (2011b): Zur Baumarteneignung bei Klimawandel: Ableitung der Trockenstress-Anfälligkeit wichtiger Waldbaumarten aus Jahrringbreiten. Environ Sci Policy, 14, 100-110.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autor


