Trockentoleranz alternativer Baumarten auf Versuchsflächen - LWF aktuell 135
von Markus Schmidt, Viviana Horna, Sharath Paligi, Bernhard Schuldt, Reiner Zimmermann, Gregor Aas
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Standort Schmellenhof (Oberfranken) des Langzeitversuches KLIP18. Im Vordergrund eine Pflanzung von Tsuga heterophylla, etwa10-jährig (© V. Horna, Universität Bayreuth)
Als Reaktion auf die Auswirkungen des Klimawandels werden vermehrt mutmaßlich trockenheitstolerante nichtheimische Baumarten im Waldbau gefördert. Ihr ökophysiologisches Verhalten unter den hiesigen Standortsbedingungen ist wenig bekannt. Deshalb ist es wichtig, neben der Leistung solcher Arten auch ihre Anpassungsfähigkeit und Stresstoleranz in unseren Wäldern zu ermitteln.
In einem Pilotprojekt unter Leitung der Universität Bayreuth wurden sechs alternative und eine heimische Baumart im Kulturstadium hinsichtlich ihrer Trockentoleranz untersucht. Dies erfolgte auf den beiden bayerischen Flächen des Langzeitversuchs KLIP18 (vgl. Frischbier et al. 2019) mittels feldtauglicher Methoden zur Quantifizierung ökophysiologischer und hydraulischer Kenngrößen und zur kontinuierlichen Erfassung des Stammdickenzuwachses. Das KLIP18-Experiment bot die Möglichkeit, dies an alternativen Baumarten identischer Herkünfte in einer für Mitteleuropa typischen Matrix von Klima- und Bodeneigenschaften zu testen. Im Rahmen von KLIP18 wurden im Jahr 2012 Versuchspflanzungen für einen Beobachtungszeitraum von 50 Jahren mit folgenden Baumarten angelegt:
- Orient-Buche (Fagus orientalis) aus dem südöstlichen Balkan
- Silber-Linde (Tilia tomentosa), beheimatet in Südosteuropa und in Kleinasien
- Libanon-Zeder (Cedrus libani) aus dem Taurus (Türkei)
- Westliche Hemlocktanne (Tsuga heterophylla) aus dem kühl-gemäßigten pazifischen Nordamerika
- Riesen-Lebensbaum (Thuja plicata), ebenfalls aus dem westlichen Nordamerika
- Bornmüller-Tanne (Abies bornmuelleriana), heimisch in den Bergen der Nordwest-Türkei
- Stiel-Eiche (Quercus robur) als einheimische Referenzbaumart
Bayerische KLIP18-Flächen
Die zwei Versuchsflächen liegen in Nordbayern. Die unterfränkische Fläche im Maintal bei Großostheim (125 m NN) zeichnet sich durch geringere Niederschläge und höhere Temperaturen sowie sandigen Boden (Flugsand, > 2 m Mächtigkeit) aus. Die oberfränkische Versuchsfläche bei Schmellenhof (Abbildung 1) liegt auf 495 m NN und hat ein eher kühl-feuchtes Klima sowie einen stark wechselfeuchten, tonig-lehmigen Boden mit wasserstauenden Schichten in circa 0,5–0,8 m Tiefe (Pseudogley).
Während der Vegetationsperioden der Jahre 2020 und 2021 wurden an jeweils fünf Individuen aller sieben Baumarten das Stammdickenwachstum halbstündlich erfasst und Daten zum Mikroklima sowie zur Bodenfeuchtigkeit in zwei Tiefen erhoben. In mehreren Freiland-Messkampagnen bestimmte man unter anderem das Blatt-Wasserpotential und im Labor die hydraulische Leitfähigkeit des Xylems.
Witterung 2020 und 2021
Großostheim war in beiden Jahren die wärmere und trockenere Fläche (Abbildung 2). Von Mai bis September 2020 war es dort mit 17,6 °C im Mittel um 2,5 °C wärmer als auf der Fläche bei Schmellenhof, im selben Zeitraum 2021 mit 16,8 °C um 2,2 °C. Die relative Luftfeuchtigkeit war in beiden Jahren von Mai bis September in Großostheim 10 % niedriger als in Schmellenhof, die Niederschlagssumme um 80 mm geringer. Von Oktober 2020 bis April 2021 fielen in Großostheim mit 285 mm etwa 70 mm weniger Niederschlag als in Schmellenhof.
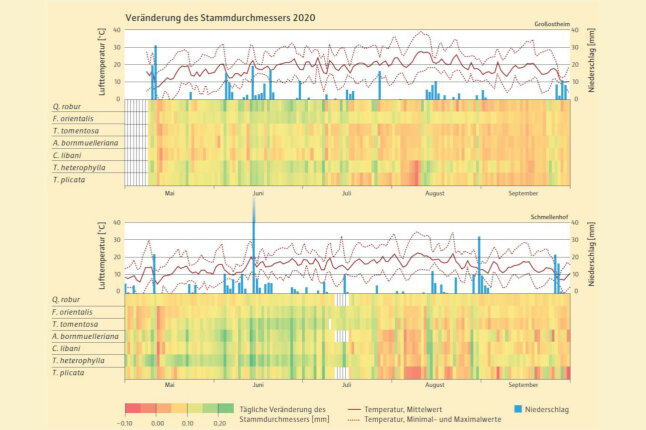
Abb. 2: Unterschiedliche Farben zeigen die tägliche Veränderung des Stammdurchmessers bei den gemessenen Tagesniederschlägen und -temperaturen. Die durchgehende rote Linie stellt den Mittelwert der Temperaturen dar, die gepunktete Linie Maximal- bzw. Minimalwerte. (© LWF)
Messungen der Stammdickenänderungen
Hochaufgelöste Messungen der Stammdickenänderung zeigen die unmittelbare Reaktion der Bäume auf Veränderungen der mikroklimatischen Bedingungen. Sie geben Hinweise auf den Wasserzustand und das (Netto-)Wachstum von Bäumen (Abbildung 2).
2020 war durch das Auftreten längerer Trockenphasen während der Vegetationsperiode charakterisiert. Eine Trockenphase im August in Verbindung mit hohen Temperaturen führte bei den meisten Arten an beiden Standorten zu einem insgesamt geringeren Stammzuwachs als im Juni und Juli. Nach starken Regenfällen nahmen alle Baumarten ausreichend Bodenwasser auf und konnten ihre Stammgewebe aufsättigen, sichtbar im Anschwellen der Stämme. Im Jahr 2021 traten keine längeren Trockenphasen auf und es konnten keine durch Trockenstress bedingten Durchmesserreaktionen festgestellt werden. Daher war das Stammwachstum 2021 größer als 2020. Eine interessante Beobachtung zu Beginn der Messungen im Jahr 2020 war die sichtbare Schrumpfung des Stammdurchmessers aller Arten während eines starken Spätfrostes Mitte Mai.
Zwischen den beiden Standorten wurden Unterschiede im Stammzuwachs beobachtet. So reagierte z.B. die Silber-Linde in Großostheim empfindlich gegenüber hohen Temperaturen und Bodentrockenheit. Sie hatte dort den geringsten Zuwachs aller Arten, während sie in Schmellenhof durchaus gut zuwuchs (Abbildung 3). Den maximalen Dickenzuwachs aller untersuchten Arten zeigte die Hemlocktanne in beiden Jahren in Schmellenhof mit 6,9 mm 2020 und 8,8 mm 2021. Bei den anderen Arten lag der Stammzuwachs bei 3–5 mm und unterschied sich nicht signifikant zwischen beiden Standorten.
Abb. 3: Mittlerer Anfangsdurchmesser, Dauer der Wachstumsperiode und Stammdickenzuwachs (n = 5/Art) im Jahr 2021 © LWF| Baumart | Großostheim 2021
Anfangs Ø [cm] | Großostheim 2021
Wachstum
[Tage] | Großostheim 2021
Dickenzuwachs
[mm] | Schmellenhof 2021
Anfangs Ø [cm] | Schmellenhof 2021
Wachstum
[Tage] | Schmellenhof 2021
Dickenzuwachs
[mm] |
|---|
| Q. robur | 3,8
± 0,8 | 113 | 4,07
± 0,89 | 5,1
± 1,7 | 111 | 5,07
± 1,69 |
|---|
| F. orientalis | 5,3
± 1,0 | 124 | 4,95
± 1,15 | 4,8
± 1,4 | 124 | 4,79
± 1,37 |
|---|
| T. tomentosa | 2,1
± 0,4 | 81 | 2,32
± 0,60 | 4,9
± 0,7 | 129 | 4,97
± 0,75 |
|---|
| A. bormuelleriana | 3,7
± 1,2 | 136 | 3,57
± 1,10 | 4,1
± 0,9 | 134 | 4,13
± 0,93 |
|---|
| C. libani | 3,2
± 1,3 | 136 | 3,33
± 1,23 | 4,7
± 0,7 | 136 | 4,71
± 0,68
|
|---|
| T. heterophylla | 7,1
± 0,7 | 151 | 6,55
± 1,29 | 8,8
± 1.7 | 168 | 8,83
± 1,75 |
|---|
| T. plicata | 5,5
± 0,9 | 144 | 5,06
± 1,27 | 4,4
± 0.8 | 116 | 4,38
± 0,81 |
|---|
Saisonalität des Zuwachses
Anhand der aus den Dendrometerdaten ermittelten effektiven Dickenzuwächse wurde die Dauer der Wachstumsperiode für 2021 ermittelt. Die Messungen hatten 2020 Ende April in Schmellenhof und Anfang Mai in Großostheim begonnen, als das Dickenwachstum bei relativ warmer Witterung bereits eingesetzt hatte. In Abbildung 3 sind für das Jahr 2021 die berechneten Werte des jährlichen Stammzuwachses und die Dauer der Zuwachsperiode für alle Arten zusammengefasst. Die Wachstumsperiode begann an beiden Standorten bei Nadelbäumen früher (13. bis 21. April) als bei Laubbäumen (9. bis 13. Mai). Eine Ausnahme bildete Thuja, die erst am 26. April in Großostheim und am 3. Mai in Schmellenhof Dickenwachstum zeigte. Hemlocktanne war die Baumart, die über den längsten Zeitraum einen Stammzuwachs aufwies (168 Tage in Schmellenhof), Silber-Linde die mit dem kürzesten (81 Tage in Großostheim).
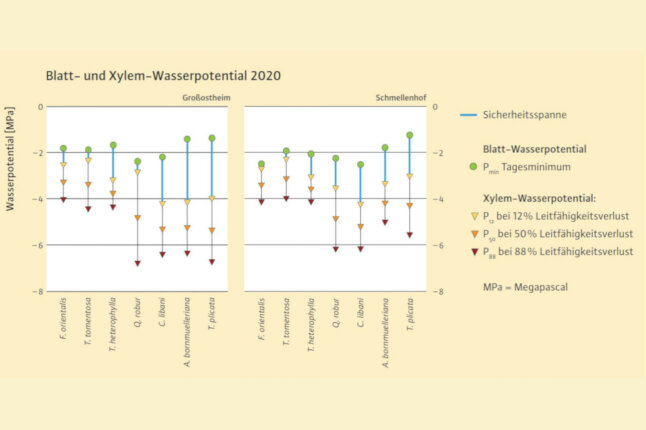
Abb. 4: Messergebnisse verschiedener hydraulischer Parameter, jeweils negativster gemessener Mittelwert für Pmin (n = 5) des Jahres 2020 beider Flächen für die jeweilige Art. Die blaue Linie entspricht der Sicherheitsspanne, hier der Abstand zwischen Pmin und P12 (Maß für den Sicherheitspuffer zur beginnenden Emboliebildung). Pmin-Daten für T. plicata 2021 (2020 nicht gemessen). Pmin-Werte in Großostheim sind weniger negativ als in Schmellenhof, weil es kurz vor den Messungen geregnet hatte. (© LWF)
Hydraulische Grenzen der Trockentoleranz
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Am empfindlichsten auf Trockenheit reagierte die Orient-Buche (© LWF)
Das wasserleitende Gewebe im Stamm stellt die Verbindung zwischen dem Wasserreservoir im Boden und den Orten der Transpiration, den Blättern, her. Da diese Bewegung hin zu den Blättern erfolgt, steht das Wasser in den Baumstämmen unter Zugspannung (Kraft pro Fläche mit negativem Vorzeichen, also das Inverse von Druck). Die Eigenschaften dieses hydraulischen Systems sind von zentraler Bedeutung für die Trockentoleranz einer Baumart.
Bei zunehmender Bodenaustrocknung und trocken-warmer Luft gerät das Wasser in den Leitbahnen unter Zugspannung. In Abhängigkeit von der Ausgestaltung der Leitbahnen werden unterschiedlich starke Zugspannungen (negative Drücke) ausgehalten, bevor es in den Leitbahnen des Xylems zu Emboliebildung, d. h. der Entstehung von Luftblasen kommt und die Wasserfäden reißen.
Die Charakteristik des zunehmenden Leitfähigkeitsverlustes des Xylems bei steigender Zugspannung (Embolieempfindlichkeit) wird im Labor an Trieben anhand von Vulnerabilitätskurven bestimmt. An diesen Kurven wird der Xylem-Unterdruck (= Xylem-Wasserpotential) bestimmt bei einem Leitfähigkeitsverlust von:
- 12 % (P12, beginnende Emboliebildung)
- 50 % (P50, kritischer Wert für das Überleben bei Nadelbäumen: Brodribb & Cochard, 2009)
- 88 % (P88, kritischer Wert für das Überleben bei Laubbäumen: Urli et al., 2013)
Strategien der Blatt-Wasserhaushaltsregulation
Das Xylem-Wasserpotential (Zugspannung in den Xylemgefäßen) schwankt täglich zwischen einem entspannt(er)en Zustand vor Sonnenaufgang und einem mehr oder weniger gespannten Zustand (negativeres Wasserpotential) am Mittag. Dies ist auf den erhöhten Wasserbedarf durch Transpiration bei begrenzter Aufnahmefähigkeit durch die Wurzeln zurückzuführen. Je wärmer und trockener die Umgebungsluft und je geringer die Wassernachleitung aus dem Boden, desto negativere Werte erreicht das Wasserpotential in den Blättern und im Xylem. Pflanzen können die zunehmende Zugspannung jedoch durch Schließen der Spaltöffnungen begrenzen.
Diese konservative Strategie von Bäumen, sich durch frühzeitigen Spaltöffnungsschluss vor Embolien zu schützen (Isohydrie), geht auf Kosten des Wachstums, da dadurch der Eintritt von Kohlendioxid für die Photosynthese behindert wird. Ein später Spaltenschluss bei niedrigeren Wasserpotentialen (Anisohydrie) riskiert die Embolie von Leitbahnen, erhält aber die Photosynthese länger aufrecht. Die Übergänge zwischen den Strategien sind fließend, anisohydrische Arten gelten als trockentoleranter. Aus dem minimal erreichten Wasserpotential Pmin einer Art und dem ermittelten Wasserpotential, bei dem die Emboliebildung einsetzt (P12), ergibt sich eine hydraulische Sicherheitsspanne und ein Hinweis auf die Trockenheitsempfindlichkeit einer Art (Abbildung 4).
Strategien der Trockenstresstoleranz
 Zoombild vorhanden
Zoombild vorhanden
Abb. 6: Die Libanonzeder erwies sich in dem Projekt als die trockentoleranteste Baumart (© LWF)
Unsere Daten liefern erste Rückschlüsse, wie die untersuchten Baumarten auf Trockenstress reagieren. Die Orient-Buche und Silber-Linde zeigten im Vergleich zu den anderen Arten ein deutlich embolieempfindlicheres Xylem (wenig negative P12-, P50- und P88-Werte, also schnelles Erreichen der Embolieschwelle).
Die Orient-Buche reagierte im Dickenwachstum (fast) nicht auf Trockenheit und schränkte die Spaltöffnungen kaum ein (sehr negatives Pmin; anisohydrisches Verhalten), kam dadurch aber sehr nahe an die Grenze zur beginnenden Emboliebildung. Vermutlich ist daher Orient-Buche die trockensensitivste der untersuchten Arten. Bei Silber-Linde wurden eine frühe, empfindliche Reaktion der Spaltöffnungen (wenig negatives Pmin; isohydrisch), die auf Kosten des Dickenwachstums ging, und eine geringe Sicherheitsspanne festgestellt. Deshalb wird diese Art als trockensensitiv eingestuft.
Die Stiel-Eiche reagierte wie die Orient-Buche im Dickenwachstum nicht auf Trockenheit. Die Unempfindlichkeit der Stiel-Eiche gegenüber recht negativen Blattwasserpotentialen (anisohydrisch) in Kombination mit einer geringen Embolieempfindlichkeit und einer sehr großen Sicherheitsspanne deuten darauf hin, dass die Art trockenheitstolerant ist.
Von den Nadelbaumarten wies die Hemlocktanne, ähnlich wie Orient-Buche und Silber-Linde, eine hohe Embolieempfindlichkeit und das stärkste Wachstum aller Arten auf. Ihr Wachstum reagierte aber sensitiv auf Trockenheit, ebenso wie ihre Spaltöffnungen. Die Sicherheitsspanne war etwas größer als bei Orient-Buche und Silber-Linde, weshalb die Hemlocktanne als etwas weniger trockensensitiv eingeordnet wird.
Die Bornmüller-Tanne reagierte im Zuwachs empfindlich auf Trockenheit, ebenso im Blattwasserpotential, bei einer gleichzeitig sehr großen Sicherheitsspanne. Bei dieser Art und beim Riesen-Lebensbaum wurde eine plastische Anpassung des hydraulischen Systems an den trockenen Standort in Großostheim festgestellt. Dies erlaubt möglicherweise ein längeres Ausharren bei Trockenheit. Fehlende Pmin-Werte des Riesenlebensbaums für 2020 lassen lediglich eine grobe Einordnung zu. Aufgrund des ähnlichen Verhaltens wie die Bornmüller-Tanne können beide Arten als bedingt trockensensitiv eingestuft werden. Die Libanon-Zeder zeigt mit sehr geringer Embolieempfindlichkeit, sehr großer Sicherheitsspanne bei niedrigem Pmin (anisohydrisch) und kaum durch Trockenheit beeinträchtigtem Wachstum die deutlichste Anpassung an trockene Bedingungen.
Daraus ergibt sich eine erste Einschätzung der Arten in Bezug auf ihre Trockenempfindlichkeit in folgender Reihenfolge:
Orient-Buche > Silber-Linde > Hemlocktanne > Bornmüller-Tanne ≈ Riesen-Lebensbaum > Stiel-Eiche > Libanon-Zeder.
Die Reihung basiert auf an Jungbäumen ermittelten Ergebnissen. Bei älteren Bäumen kommen weitere Faktoren hinzu, z. B. die Wasserspeicherkapazität im Stamm, die Durchwurzelungstiefe oder die Kronen- und Wurzelkonkurrenz im Bestand.
Ausblick
Die eingesetzten Methoden zur Erfassung der Reaktionen einzelner Baumarten auf die Variabilität von Klima, Witterung und Boden geben einen Einblick in Anpassungsmöglichkeiten an Trockenheit. Das Ergebnis ist ein Mosaik von Eigenschaften, das noch Lücken aufweist. So konnten in der kurzen Zeit dieser Pilotstudie nur einige der Merkmale untersucht werden, die bei Pflanzen die Trockentoleranz bedingen. Auch wies die Witterung während des Untersuchungszeitraums nur wenig extreme Trockenperioden auf, wie sie 2018 und 2019 über längere Zeit herrschten. Dennoch erwiesen sich alle Methoden als geeignet, die unmittelbare ökophysiologische Reaktion der Bäume auf die Variabilität der Witterung zu analysieren. So wurden beispielsweise auch die Auswirkungen eines starken Spätfrostereignisses mit den Dendrometern erfasst. Die Spätfrostempfindlichkeit ist ein weiterer wichtiger Faktor bei der Auswahl alternativer Baumarten. Spätfröste stellen zukünftig ein großes Problem auf wieder zu bewaldenden Schadflächen dar, da Jungbäume auf Freiflächen starker nächtlicher Auskühlung (Strahlungsfrost) ausgesetzt sein können. Die angewandten Methoden bieten sich für weitere Untersuchungen sowohl im Rahmen Monitoring-basierter Ansätze als auch für experimentelle Austrocknungs- und Bewässerungsversuche an.
Zusammenfassung
Ziel des Pilotprojekts ist die Beurteilung der Trockensensitivität von Jungpflanzen alternativer Baumarten anhand ökophysiologischer und hydraulischer Kenngrößen. Mit Hilfe der angewandten Methoden wurden 2020 und 2021 erste Erkenntnisse über die artspezifischen ökophysiologischen Reaktionen auf Unterschiede in der Witterung und den Standort gewonnen. Die Methoden bieten sich für weitere Untersuchungen sowohl im Rahmen Monitoring-basierter Ansätze als auch für experimentelle Austrocknungs- und Bewässerungsversuche an.
Das Projekt »Potential alternativer Baumarten im Klimawandel - Früherkennung von Trockenstress auf neuen Versuchsflächen in Bayern« (klifW002) (Laufzeit: 01.05.2020 - 31.12.2021) wurde vom Bayerischen Staatsministerium für Ernährung, Landwirtschaft und Forsten finanziert, ebenso die Anlage des Langzeitversuches KLIP18 (Laufzeit: 01.02.2009 - 31.12.2012). Seit Ende der Laufzeit wird KLIP18 von der Bayerischen Landesanstalt für Wald und Forstwirtschaft finanziert.
Literatur
- Brodribb, T.J.; Cochard, H. (2009): Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiology 149, 575-584.
- Frischbier, N.; Nikolova, P.S.; Brang, P.; Klumpp, R.; Aas, G.; Binder, F. (2019): Climate change adaptation with non-native tree species in Central European forests: early tree survival in a multi-site field trial. European Journal of Forest Research 138, 1015–1032.
- Urli, M.; Porté, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. (2013): Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiology 33, 672–683.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren



 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
