


Andreas König
Artgerechte Überwinterung und Notzeit von Rehen – LWF aktuell 129
AG »Wildbiologie und Wildtiermanagement« der TUM forscht zum Thema Winterfütterung – mit aufschlussreichen Ergebnissen
Alle Jahre wieder erscheint das Thema Winterfütterung, sobald es etwas winterlich mit Schnee wird. Diskussionen, Forderungen und Aussagen sind in der Regel emotional. Daher lohnt es, wissenschaftlichen Fakten zur Energieversorgung der Rehe durch die Vegetation im Herbst und Winter, ihren Umgang damit und ihre aufgenommene Energie sowie der Frage, wann eigentlich Notzeit sein könnte, nachzugehen.
Diesen Aufgaben widmet sich seit einigen Jahren die AG »Wildbiologie und Wildtiermanagement« am Lehrstuhl für Tierernährung der TU München.
Über wen reden wir?
Wann kann es theoretisch zu einer Notzeit für Rehe kommen?
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: : Schematische Darstellung der Zusammenhänge zwischen Energieaufnahme, Fettreserven und Energiebedarf bei Rehen im Jahresverlauf (Grafik: LWF)
Abbildung 1 zeigt den Zusammenhang zwischen Nahrungsaufnahme, Fettreserven und Energiebedarf bei Rehen im Jahresverlauf (Hofmann 1981). Notzeit gibt es bei Wildtieren dann, wenn alle Reserven abgebaut sind, der Stoffwechsel steigt und die Nahrung keine Energie liefert. Nach Hofmann (1981) kann das bei Wildwiederkäuern nur im März und April sein, hohe Schneelagen im Januar und Februar spielen keine Rolle (König & Zannantonio 2006). Das deckt sich zum Beispiel mit den höchsten Rissraten von Wölfen im März/April (Smith et al. 2004), da ihre Beutetiere geschwächt sind. Dann bringen auch Bartgeier ihre Jungen synchron mit dem hohen Fallwildaufkommen zur Welt.
In den letzten 40 Jahren haben sich die phänologischen Phasen durch den Klimawandel verschoben und die Vegetation beginnt heute gut 15 Tage früher zu ergrünen (Menzel 2006; Stahl 2016). So fällt der April als mögliche Notzeit aus. Aktuell könnte es daher, wenn überhaupt, nur im März zu einem Engpass und einer möglichen Notzeit für Wildwiederkäuer kommen.
Fettreserven und Körpermasse der Rehe im Jahresverlauf
 Zoombild vorhanden
Zoombild vorhanden
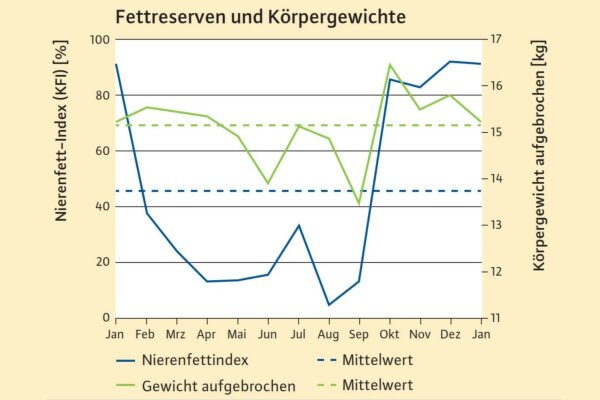
Abb. 2: Nierenfettindex (blau) als Maß für die Fettreserven (KFI, N=234) und Körpergewicht, aufgebrochen (grün) von einjährigen und mehrjährigen Rehen (N=173) Grafik: LWF)
Wie bei Hofmann (1981) beschrieben (Abbildung 1), legen Rehe im Herbst und Frühwinter hohe Fettreserven an, um niedrigere Energiedichten der Vegetation im Winter auszugleichen. Ab Februar beginnen sie, diese Fettreserven abzubauen (Abbildung 2, blaue Linie). Das bestätigen aktuelle Daten zur Rehernährung für Bayern (König et al. 2016). Weiterhin bleibt die Körpermasse zwischen Januar und April etwa konstant (grüne Linie). Laut den bayerischen Daten benötigen Rehe ihre Fettreserven im Spätwinter und Frühjahr nicht bzw. nur in geringem Umfang, sonst hätte ihre Körpermasse abgenommen.
Verluste der Körpermasse im Laufe des Winters bei Rehen werden zwischen 20 und 30 % als normal betrachtet (Bubenik 1971; Holand et al. 1998). Im Durchschnitt wiegen unsere untersuchten erwachsenen Rehe im Winter 15,5 kg und im Frühjahr 15,1 kg, was dem Jahresdurchschnitt von 15,1 kg entspricht.
In laufenden Studien zeigen adulte ungefütterte Rehe aus den Bergen mit ca. 17,2 kg die höchsten Durchschnittsgewichte.
Energiedichte der Nahrung von Rehen
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Nur weil Schnee liegt, haben die Rehe noch lange keine Notzeit. (Foto: ©PantherMedia / Lubos Chlubny)
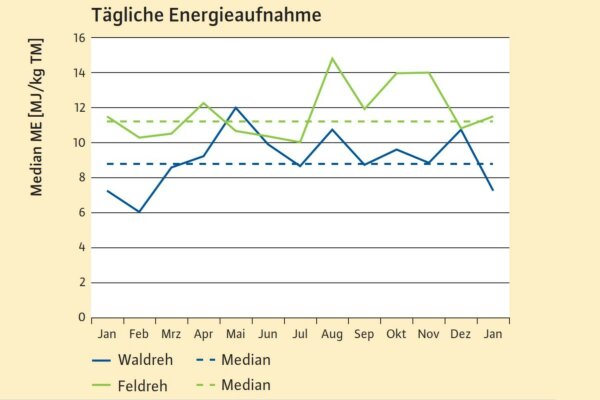
Das Argument für eine Fütterung, die Kulturlandschaft biete keine Äsung für Rehe, entbehrt somit der wissenschaftlichen Grundlage. Durch die hohe Energiedichte der Äsung im Agrargebiet im Winter und Frühjahr, die über dem Jahresdurchschnitt des Waldes liegt, unterliegen auch Feldrehe im Winter keiner Beeinträchtigung. Wie zu erwarten, sind im Januar und Februar die Energiedichten in der Naturlandschaft abgesunken, jedoch erreicht die früh einsetzende Vegetation im März das Energieniveau des Jahresdurchschnitts.
Wie gehen Rehe mit schwankender Energiedichte um?
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: : Median der Energiedichte der von Rehen aufgenommenen Äsung (N=245) (Grafik: LWF)
Mit diesen Anpassungen decken Rehe in der Regel ihren Energiebedarf von ca. 3,2–4,3 Megajoule (MJ) pro Reh und Tag das ganze Jahr aus der vorhandenen Äsung (Bobek 1980; Hartfiel et al. 1985; Onderscheka 1999; Oslage & Strothmann,1988; Weiner 1977).
Beschaffenheit der Äsung unserer Rehe
 Zoombild vorhanden
Zoombild vorhanden
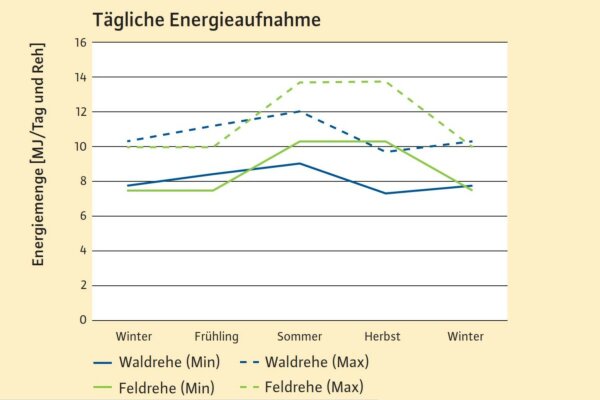
Abb. 5: Median der pro Tag und Reh aufgenommenen umsetzbaren Energie, differenziert für Waldrehe und Feldrehe (N=245) (Grafik: LWF)
Eine so enge Zuordnung zwischen den Äsungstypen und den durch die Arten aufgenommen Pflanzen wie von Hofmann (1982 & 1989) beschrieben, besteht nicht (Behrend et al. 2004; Clauss 2010; Djordjevic et al. 2006; Hartfiel et al. 1985; Holand 1994; Lechner et al. 2009; Obidzinski et al. 2017; Robbins et al. 1995; Serrano Ferron et al. 2011; Woodall 1992). Im Gegenteil:
In der Rehäsung finden sich im Durchschnitt nicht nur Rohfaseranteile von 21–38 % mit Maxima von 49 % Trockenmasse, die jenen von Rotwild (25–30 %) oder Mufflon (22–34 %) (Briedermann et al. 1988; Drescher-Kaden & Seifelnasr 1977) entsprechen, sondern sie zeigen auch eine üppige Ausstattung ihres Pansenmikrobioms mit faserzersetzenden Bakterien und Pilzen (Dahl et al. 2020; König et al. 2016).
Die Notzeit-Mast
In der aktuellen Fütterungspraxis werden als Notzeitfutter vielfach Mischungen mit 30–50 % Getreideanteilen (z. B. Hafer, Gerste, Mais) und entsprechend wenig Raufutter (z. B. Grummet) empfohlen, welche durch deutlich zu niedrige Faseranteile bei gleichzeitig zu hohen Protein- und Kohlenhydratanteilen gekennzeichnet sind.
Derartige Mischungen enthalten ca. 15 % Fasern pro Kilogramm Trockenmasse, und das in einer Jahreszeit, in der Rehe freiwillig zwischen 26 und 49 % Trockenmasse Fasern aufnehmen. So niedrige Faseranteile bei gleichzeitig hohem Protein- und Kohlehydratangebot im Pansen führen zur Bildung von Propion- und Milchsäure, der Pansen pH-Wert sinkt unter pH 6, wodurch eine Pansenazidose verursacht wird (Deutz et al 2009). Pansenazidose ist in Österreich der Hauptverursacher für Fallwild bei Rehen im Winter (Arnold 2020).
Rehe mit zu niedrigem pH-Wert im Pansen nehmen aus der Waldvegetation Fasern auf, um ihren pH-Wert anzuheben. Diese Futtermischungen weisen eine Energiedichte von ca. 10– 12 Megajoule umsetzbare Energie (ME MJ) pro Kilogramm Trockenmasse (TM) auf, während natürliche Äsung eine Energiedichte zwischen 4 und 6,3 ME MJ/kg TM bereitstellt.
Diese vorgeschlagene Notzeitfütterung hat eine Energiedichte, die normal für »Hochleistungsmilchkühe« und zur »Mast« verwendet wird. Getreide jeglicher Art bewirkt zu hohe Energiedichten und ist kein Erhaltungsfutter im Sinne einer Notzeitfütterung. Im Gegenteil –Getreide in einer Fütterung zwingt das Reh regelrecht dazu, verstärkt Fasern aufzunehmen, um den pH-Wert im Pansen anzuheben. Damit trägt Getreide in der Fütterung zu weiterem Verbiss bei.
Zusammenfassung
Energiedefizit und Notlage könnten sie höchstens im März haben, was auf Grund des Klimawandels aber sehr unwahrscheinlich ist. Schnee und Frost bedeuten dagegen per se keine Notzeit. Potenzielle Notzeit muss immer revierweise unter Berücksichtigung der Kondition der Tiere und der vorhandenen natürlichen Äsung beurteilt werden. Häufig vorgeschlagene Futtermischungen haben nicht nur zu wenig Faseranteile für Rehe, sondern haben eine Energiedichte, die in der Regel zur Mast von landwirtschaftlichen Nutztieren verwendet wird. Getreide jeder Art hat in einer Erhaltungsfütterung nichts zu suchen.
- Andersen, R.; Duncan, P.; Linnell, J. D. C. (Eds.) (1998): The European Roe Deer: The Biology of Success. Oslo: Scandinavian University Press
- Arnold, W. (2003): Neue Erkenntnisse zur Winterökologie des Rotwildes - der verborgene Winterschlaf. In Jagdverband, B. (Ed.), Rotwild in Bayern - heute und morgen; S. 15–20
- Arnold, W. (2013): Jahreszeitliche Anpassung bei Wildwiederkäuern - wo steht das Rehwild? In BAYERN, L. (Series Ed.), Schriftenreihe des Landesjagdverbandes: Vol. 20. Hege und Bejagung des Rehwildes; S. 13–21
- Arnold, W. (2020): Jahresbericht 2019. In: Jahresberichte. Wien: Forschungsinstitut für Wildtierkunde und Ökologie
- Behrend, A.; Lechner-Doll, M.; Streich, W. J.; Clauss, M. (2004): Seasonal faecal excretion, gut fill, liquid and particle marker retention in mouflon ovis ammon musimon, and a comparison with roe deer. Acta Theriologica, 49(4), S. 503–515
- Bobek, B. (1980): A model for optimization of roe deer management in Central Europe. Journal of Wildlife Management, 44(4), S. 837–848
- Briedermann, L.; Dittrich, G.; Lockow, K.-W. (1988): Rotwild Cervus elaphus L. In: Stubbe, H. (Ed.), Buch der Hege Band 1 Haarwild (S. 2–57). Thun, Frankfurt a.M.: Verlag Harri Deutsch
- Bubenik, A. B. (1971): Rehwildhege und Rehwildbiologie. München: F.C. Mayer Verlag
- Bubenik, A. B. (1984): Ernährung, Verhalten und Umwelt des Schalenwildes. München, Wien, Zürich: BLV Verlagsgesellschaft
- Clauss, M. (2010): Artgerechte Fütterung der Wildtiere - was ist fachlich vertretbar. Paper presented at the Östereichische Jägertagung, Raumburg-Gumpenstein
- Dahl, S.-A.; Hudler, M.; Windisch, W.; Bolduan, C.; Brugger, D.; König, A. (2020): High fibre selection by roe deer (Capreolus capreolus): evidence of ruminal microbiome adaption to seasonal and geographical differences in nutrient composition. Animal Production Science. doi:10.1071/an19376
- Deutz, A.; Gasteiner, J.; Buchgraber, K. (2009): Fütterung von Reh- und Rotwild. Ein Praxisratgeber. Leopold Stocker Verlag
- Djordjevic, N.; Popovic, Z.; Grubic, G. (2006): Chemical composition of the rumen contents in roe deer (Capreolus capreolus) as potetial quality indicator of their feeding. Journal of Agricultural Sciences, 51(2), S. 133–140
- Drescher-Kaden, U.; Seifelnasr, E. A. (1977): Untersuchungen zum Verdauungstrakt von Rehwild, Damwild und Mufflon. Mitteilung 2: Rohnährstoffe im Panseninhalt von Reh, Damhirsch und Mufflon. Z. Jagdwiss., 23, S. 6–11
- Geist, V. (1999): Deer of the world. Their Evolution, Behaviour, and ecology. Shrewsbury: Swan Hill Press
- Hartfiel, W.; Pheiffer, J.; Dissen, J. (1985): Energetische Untersuchungen an Reh und Schaf mit Hilfe der quantitativen Thermographie zur Beurteilung des Energiebedarfs im Winter. Z. Jagdwiss., 31, S. 34–41
- Hofmann, A.; Nievergelt, B. (1972): Das jahreszeitliche Verteilungsmuster und der Äsungsdruck von Alpensteinbock, Gemse, Rothirsch und Reh in einem begrenzten Gebiet im Oberengadin. Z. Jagdwiss., 18, S. 185–212
- Hofmann, R. R. (1981): Über die Notzeit des Schalenwildes in der Kulturlandschaft - wissenschaftliche Erkenntnisse, gesetzliche Bestimmungen und Hegepraxis. In: Hofmann (Ed.), Wildbiologische Informationen für den Jäger (Vol. IV). Stuttgart: Ferdinand Enke Verlag
- Hofmann, R. R. (1982): Die Stellung der europäischen Wildwiederkäuer im System der Äsungstypen. In: Hofmann (Ed.), Wildbiologische Informationen für den Jäger. Jagd + Hege - Ausbildungsbuch I (S. 9-18). Stuttgart: Ferdinand Enke Verlag.
- Hofmann, R. R. (1989): Evolutionary steps of ecophysiological adaption and diversification of ruminants: a comperative view if their digestives system. Oecologia, 78(4), S. 443–457
- Holand, O. (1994): Seasonal dynamics of digestion in relation to diet quality and intake in European roe deer (Capreolus capreolus). Oecologia, 98, S. 274–279
- Holand, O.; Mysterud, A.; Wannang, A.; Linnell, J. D. C. (1998): Roe deer in northern environments: Physiology and behaviour. In: Andersen, Duncan, Linnell (Eds.), The European Roe Deer: The Biology of Success (S. 117–138). Oslo: Scandinavian University Press
- König, A.; Zannantonio, D. (2006): Gamswildmonitoring 1998–2006 im Modellprojekt Nationalparkreviere Gasteinertal
- König, A.; Scheingraber, M.; Mitschke, J. (2016): Energiegehalt und Qualität der Nahrung von Rehen (Capreolus capreolus) im Jahresverlauf in zwei unterschiedlich geprägten Habitaten. In: Zentrum-Wald-Forst-Holz (Series Ed.), Forstliche Forschungsberichte (S. 199), Freising
- König, A.; Hudler, M.; Dahl, S.-A.; Bolduan, C.; Brugger, D.; Windisch, W. (2020): Response of roe deer (Capreolus capreolus) to seasonal and local changes in dietary energy content and quality. Animal Production Science. doi:10.1071/an19375
- Lechner, I.; Barboza, P.; Collins, W.; Günther, D.; Hattendorf, B.; Hummel, J.; Clauss, M. (2009): No ‚bypass‘ in adult ruminants: Passage of fluid ingested vs. fluid inserted into the rumen in fistulates muskoxen (Ovisbos moschatus), reeindeer (Rangifer tarandus) and moose (Alces alces). Comperative Biochemistry and Physiology Part A, 154, S. 151–156
- Leonhardt, P. (1986): Jagdrecht. Bundesjagdgeetz, Bayerisches Jagdgesetz, Ergänzende Bestimmungen. München: Carl Link Verlag
- Menzel, A. (2006): Zeitliche Verschiebung von Austrieb, Blüte, Fruchtreife und Blattverfärbung im Zuge der rezenten Klimaerwärmung. Forum für Wissen, S. 47–53
- Müller, F. (1982): Das Rehwild. In: Wildbiologische Informationen für den Jäger (S. 165–176). Stuttgart: Ferdinand Enke Verlag
- Obidzinski, A.; Miltko, R.; Bolibok, L.; Wajdzik, M.; Skubis, J.; Nasiadka, P. (2017): Variation of natural diet of free ranging mouflon affects their ruminal protozoa composition. Smaöö Ruminant Research, 157, S. 57–64
- Onderscheka, K. (1999): Das Rehwild - seine Ernährung und Fütterung. Paper presented at the Rehwild in der Kulturlandschaft, Nürnberg
- Oslage, H. J.; Strothmann, A. (1988): Zum Energie- und Proteinbedarf von Rehwild. Z. Jagdwiss., 34, S. 164–181
- Robbins, C. T.; Spalinger, D. E.; Hoven, W. v. (1995): Adaptation of Ruminants to browse and grass diets: are anatomical based browser-grazer interpretations vaild? Oecologia, 103, S. 208–213
- Sempere, A. J.; Sokolov, V. E.; Danilkin, A. A. (1996): Capreolus capreolus. Mammalian Species, 538, S. 1–9
- Serrano Ferron, E.; Verheyden, H.; Hummel, J.; Cargnelutti, B.; Lourtet, B.; Merlet, J.; González-Candela, M.; Angibault, J. M.; Hewison, A. J. M.; Clauss, M. (2011): Digestive plasticity as a response to woodland fragmentation in roe deer. Ecological Research, 27(1), S. 77–82. doi:10.1007/s11284-011-0872-x
- Smith, D. W.; Drummer, T. D.; Murphey, K. M.; Guernsey, D. S.; Evans, S. B. (2004): Winter prey selection and estimation of wolf kill rates in Yellowstone National Park, 1995-2000. Journal of Wildlife Management, 68(1), S. 153–166
- Stahl, B. (2016): Die Verschiebung der phänologischen Phasen potentieller Nahrungspflanzen des Rehs (Capreolus capreolus Linnaeus) während der Setzzeit als Folge des Klimawandels und die damit verbundene zeitliche Änderung der Nahrungsquantität und -qualität. (Master Master). Technische Universität München, Freising
- Stubbe, C. (1997): Das Rehwild (The Roe Deer) (4 ed.). Berlin: Parey Buchverlag
- Weiner, J. (1977): Energy Metabolism of the Roe Deer. Acta Theriologica, 22(1), S. 3–24
- Woodall, P. F. (1992): An evolution of a rapid method for estimating digestibility. African Journal of Ecology, 30, S. 181–185