


RSS-Feed der Bay. Landesanstalt für Wald und Forstwirtschaft abonnieren
So verpassen Sie keine Neuigkeiten mehr. Unser RSS-Feed "Nachrichten der Bayerischen Landesanstalt für Wald und Forstwirtschaft" informiert Sie kostenlos über unsere aktuellen Beiträge.
Ernst Segatz
Biodiversität und waldbauliche Behandlung von Edelkastanienwäldern – LWF Wissen 81
Ist von der Baumart Edelkastanie die Rede, denkt man in Deutschland wohl zuerst an einen mehr oder weniger exotischen Baum des Weinbauklimas im Südwesten, an den herben Geruch und die weiße Pracht zur Zeit der Blüte im Frühsommer sowie an Maronen als herbstlichen Genuss zum neuen Wein.
Schon die Eigenschaften und die Verwendung des Holzes sind nur wenigen bekannt und das Wissen darüber ist meist auf das Ortstypische und Ortsübliche beschränkt. Auch stellt sich bei einer ursprünglich in Mitteleuropa nicht heimischen Art eher weniger die Frage nach ihrem Beitrag zur Biodiversität. So fehlen in der Folge im Natura 2000-Netzwerk für Deutschland Edelkastanien-Lebensraumtypen (LRT).
Biodiversität des Bodenlebens
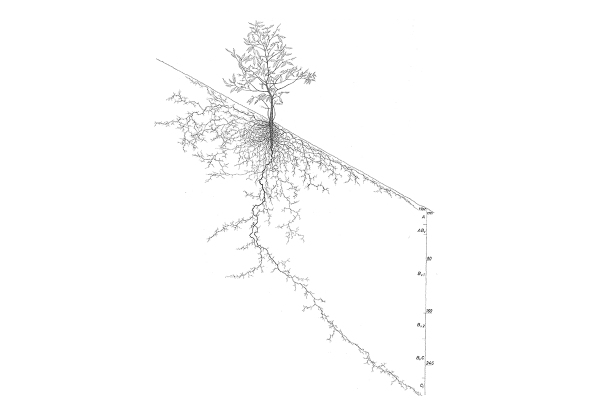 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Wurzelbild einer jungen Edelkastanie. (Quelle: Kutschera und Lichtenegger 2002)
Sie wächst optimal auf durchlässigen, sehr gut durchlüfteten Böden und als Säurezeiger in einem Bereich von pH 3,5 bis 5,5. Auf staunassen Böden reagiert die Edelkastanie empfindlich bezüglich des Wurzelwachstums und leidet dort häufig unter Befall mit [i]Phytophthora[/i]-Pilzen, die die sogenannte Tintenkrankheit hervorrufen. Deren mit Geißeln ausgestattete Sporen benötigen für die Fortbewegung im Boden freies Bodenwasser.
Die Edelkastanie wird als Tiefwurzler bezeichnet, ihre Wurzeltracht soll am ehesten jener der Eichen entsprechen (Kutschera und Lichtenegger 2002). Dies wäre eine weitere Parallele zu den [i]Quercus[/i]-Arten. Typisch sind, wie bei der Stieleiche, die Vorwüchsigkeit einer Polwurzel (Abbildung 1) und ein anfängliches Zurückbleiben der Seitenwurzeln (Kutschera und Lichtenegger 2002).
Auf tief zu durchwurzelnden Böden dringt sie bis zu den gleichmäßiger feuchten und gleichmäßiger temperierten Bodenschichten vor (Kutschera und Lichtenegger 2002). »Nach der bisher gebräuchlichen Typisierung würde das Wurzelsystem älterer Edelkastanien unter Vernachlässigung der über den Kronenrand hinausragenden Wurzeln am ehesten einem ›Herzwurzelsystem‹ entsprechen« (Kutschera und Lichtenegger 2002).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Wurzelbild eines Edelkastanien-Stockausschlags. (Quelle: Kutschera und Lichtenegger 2002)
Die Humusform unter den Edelkastanien-Beständen ist in der Regel als L- bis F-Mull anzusprechen (Abbildungen 3 und 4). Dies entspricht nicht der Erwartung, wenn man einen wie bei der Eiche hohen Gerbstoffgehalt der derben, ledrig glänzenden Blätter unterstellt. Sie sind zudem mit einer Wachsschicht überzogen, die der Selbstreinigung in Form eines »Lotus-Effekts« dient (Anders 2010). Die günstige Humusform belegt trotzdem eine leicht zersetzbare Streu.
Dies passt auch zu dem Kalium-Reichtum der Blätter (IG Edelkastanie 2006). Bodenorganismen weisen ein Kohlenstoff-Stickstoff- Verhältnis (C /N-Verhältnis) von rund 25:1 auf. Bei der Blattstreu der Edelkastanie liegt dieses mit 23:1 in einem nahezu optimalen Bereich. Nach eineinhalb Jahren sind die Blätter weitgehend zersetzt (Scheffer und Ulrich 1960). Bei der Eiche, die die Humusform Moder bildet, liegt das C /N-Verhältnis dagegen mit 47:1 deutlich ungünstiger.

Abb. 3: Blattstreu und Fruchtkapsel

Abb. 4: Blattstreu
 Zoombild vorhanden
Zoombild vorhanden
Abb 5: Streu von männlichen Edelkastanien-Blüten. (Foto: E. Segatz)
Das unter günstigen Bedingungen ausgebildete tiefgründige Herzwurzelwerk der Edelkastanie und ihre sehr gut abbaubare Streu der Blüten und Laubblätter schaffen die Voraussetzungen für ein reiches Bodenleben und eine hohe Biodiversität der Bodenorganismen.
Die Edelkastanie tritt jahreszeitlich sehr spät in die vegetative Phase ein. Austrieb und Blühphase treten normalerweise ab Mai ein, was der Edelkastanie erlaubt, vor allem während der Blüte das Risiko von Spätfrösten zu reduzieren (Conedera 2007).
Biodiversität der Bestäuber
Die Edelkastanie weist sowohl die typischen Merkmale der Insektenbestäubung als auch diejenigen der Windbestäubung auf. Beide Merkmale weisen auf ein evolutives Übergangsstadium hin (Conedera 2017). Sie entwickelt sich von einer insekten- zu einer windbestäubten Art. Die Insektenbestäubung ist üblicherweise von sekundärer Wichtigkeit, kann aber in Fällen von besonders feuchten meteorologischen Bedingungen während der Blüte wichtig sein: Der Pollen wird dann viskos, klebrig und wenig geeignet für den Transport durch die Luft. Für die Insektenbestäubung überwiegen nebst Honigbienen Käfer, Schwebfliegen und Hummelarten. Die größere Effizienz der Windbestäubung bleibt aber unbezweifelt, was der große Erfolg dieser Befruchtungsart bei trockenem und windigem Wetter beweist (Conedera 2007).
Die Kerbtiere profitieren bei der Edelkastanienblüte sowohl vom Nektar der weiblichen als auch vom Pollen der männlichen Blüten. Zeitgleich zur Nektartracht findet eine Honigtau-Tracht statt, wobei den Honigtau nach Pritsch (2007) in Schabacker, Lehnigk, Eichhorn und Hapla (2014) die Eichen-Napfschildlaus [i](Eulecanium rufulum)[/i], die Edelkastanien-Rindenlaus [i](Lachnus longipes)[/i] sowie die Edelkastanie-Zierlaus [i](Myzocallis castanicola)[/i] erzeugen.
Für den Menschen steht die Nutzung der blühenden Edelkastanienbestände als Bienenweide in der Imkerei im Vordergrund. Hier spielen nicht unbedeutende wirtschaftliche Interessen eine Rolle. Der Beitrag der Baumart zur Erhaltung der Bienen ist jedoch existentiell auch für den Menschen und kann in einer Zeit, in der das Insektensterben in aller Munde ist, nicht hoch genug gewertet werden. Dieser Beitrag wird finanziell nicht honoriert, es soll hier aber auf die Bedeutung der Edelkastanie für den Artenschutz hingewiesen werden.
Biodiversität in unterschiedlichen Bewirtschaftungsformen
Niederwald

Abb. 6: Alter Edelkastanienstock mit jungen Stockausschlägen (Foto: E. Segatz)
Niederwälder gelten als eher naturferne Standorte. Die nutzungsbedingten Eingriffe bei der Niederwaldbewirtschaftung führen jedoch zu katastrophenarti- gen Störungen innerhalb des Waldökosystems, wie sie auch in überalterten und durch natürliche Katastrophen gestörten Naturwäldern auftreten (Scherzinger 1996; Kaule 1991).
Die daraus resultierenden ökologischen Besonderheiten des Niederwalds (hoher Wärme- und Lichtgenuss, kleinklimatische Extreme, Erosionsanfälligkeit des Standorts mit Entstehung besonderer Kleinstrukturen u. a.) und dessen damit zusammenhängende Bedeutung für die Artenvielfalt treffen natürlich auch für den Edelkastanien-Niederwald zu. Da die einzelne Teilfläche in sich sehr homogen sein kann, ist eine mosaikartige Verteilung, wie dies früher beim Flächenfachwerk der Fall war, besonders günstig zu beurteilen.
Durch das sehr stark ausgeprägte Stockausschlagvermögen und das üppige Austreiben großblättriger Reiser währt die hinsichtlich der Artenvielfalt wichtigste Lichtwald-Phase mit stärkerer Besonnung auch des Waldbodens jedoch nur wenige Jahre. Selbst starker Wildverbiss hindert in der Regel die intensive Wiederbestockung und das Dunkelwerden der Jungbestände nicht entscheidend.
Große Bedeutung, insbesondere in Niederwäldern, die bereits mehrfach auf den Stock gesetzt wurden, kommt den häufig voluminösen Stöcken zu. Sie bilden einen großen Vorrat an ober- und unterirdischem Totholz in Verbindung mit der Entstehung neuen Wurzelgewebes (siehe Abbildungen 2 und 6). Der Wechsel von feucht und trocken, absterbenden Teilen und Neubildung in Kontakt mit dem Mineralboden bieten optimale Lebensbedingungen für viele Pilze und Insekten.
Hinsichtlich der Alterungs-, Absterbe- und Erneuerungsprozesse am Edelkastanienwurzelstock und deren Bedeutung für viele Organismengruppen besteht noch Forschungsbedarf.
Überalterter Niederwald, mit und ohne Eingriffe
Benner (2010) zählte rund 4.230 Stämme in einem 27 Jahre alten Edelkastanien-Bestand, davon waren 1.950 Stämme (etwas mehr als 45 %) bereits abgestorben.
Auch diese Strukturen liefern einen positiven Beitrag zur Biodiversität. Waldbaulich und ertragskundlich begründete steuernde Eingriffe zur Vereinzelung und Förderung der Wertträger wirken logischerweise diesem Prozess entgegen.
Mittelwald
 Zoombild vorhanden
Zoombild vorhanden
Abb. 7: Schaffung mittelwaldartiger Strukturen. (Foto: E. Segatz)
Mittelwaldstrukturen verbinden Elemente des Niederwalds mit den Eigenschaften solitärer Altbäume. Ein Nutzungsverzicht einzelner alter Bäume würde einen wesentlichen Beitrag zur Biodiversität liefern.
Zielstärkenorientierte, hochwaldähnliche Behandlung
Da die Edelkastanie in der Lage ist, auf guten Standorten jährlich einen Durchmesserzuwachs von rund 1 cm zu leisten, kann sie beispielsweise mit 60 Jahren bereits einen Durchmesser von 60 cm erreichen. In Reinform betrieben, bleiben auf der Fläche außer den Z-Bäume keine weiteren Bäume stärkerer Dimensionen erhalten.
Um ökologische Aspekte zu berücksichtigen, müssten hier bewusst Teilflächen von dieser Behandlung ausgenommen bzw. »ökologische Z-Bäume« mit Biotopstrukturen ausgewählt werden, die in der Regel diametral zur ökonomischen Nutzung stehen.
Erhalt von Altbeständen und Biotopstrukturen
Höhlenbäume
 Zoombild vorhanden
Zoombild vorhanden
Abb. 8: Höhlen des Buntspechts. (Foto: E. Segatz)
Totholz
Altbäume (»Methusalembäume«)
Individuen oder Bestände seltener heimischer Baumarten
Bäume mit besonderen Merkmalen
Bäume mit sich lösender Rinde oder Rindentaschen
»Obligatorische Biotopbäume«
 Zoombild vorhanden
Zoombild vorhanden
Abb. 9: Großhöhle am Stammfuß. (Foto: E. Segatz)
Edelkastanien sind, auch im Verhältnis zu den Eichen, überproportional stark hinsichtlich Großhöhlen ausgestattet, was sehr gut mit dem Entstehen der Bäume aus Stockausschlag und den damit verbundenen früheren Stammverletzungen sowie mit den Folgen von Ringschäle zu erklären ist (Abbildungen 9 und 10). Einzelne Edelkastanien mit Großhöhlen sind legendär, so wie die »Kastanie der 100 Pferde« auf Sizilien, ein Baum, in dessen Höhlung Vieh eingetrieben werden konnte. Es ist jedoch unklar, ob es sich nicht ursprünglich um mehrere zusammengewachsene Bäume gehandelt hat.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 10: Inneres einer hohlen Edelkastanie. (Foto: E. Segatz)
Wegen des Mangels an Altbäumen leider kaum zu beobachten!
Bäume mit bekannten Fortpflanzungs- und Ruhestätten von FFH-Anhang-IV-Arten (z. B. Eremit, Heldbock) und in FFH-Gebieten Anhang II Arten mit geringem Aktionsradius (z. B Veilchenblauer Wurzelhalsschnellkäfer).
Sehr alte und dicke Edelkastanien besitzen das Potenzial zu Fortpflanzungs- und Ruhestätten sehr seltener Arten. Am Niederrhein in Nordrhein-Westfalen wurde in der Edelkastanienallee bei Schloss Dyck, wahrscheinlich im Jahr 1811 gepflanzt (zum Fundzeitpunkt rund 200 Jahre alt), erstmals das Vorkommen des Eremits an Edelkastanien belegt. Köhler (2016) beobachtete bei seiner Untersuchung in der Pfalz zwar keine FFH-Arten, fand am ältesten Standort jedoch Fraßspuren des Heldbocks [i](Cerambyx cerdo)[/i]. Er geht davon aus, dass sich auch die anderen aus Rheinland- Pfalz bekannten FFH-Totholzkäferarten an Edelkastanie entwickeln können.
Edelkastanienstöcke sind als Substrat für die Larven des Hirschkäfers geeignet (Rinck 2018, mdl. Mitt.).
Über den Schutz von Einzelbäumen hinaus geht die Ausweisung von Biotopbaumgruppen und flächig definierter Elemente sogenannter Waldrefugien. Infolge von Rindenkrebs oder [i]Phytophthora[/i] abgestorbene Baumgruppen könnten unter Berücksichtigung der Verkehrssicherungspflicht als ökologisch wertvolle Kleinflächen erhalten werden, insbesondere auf ungünstigeren Standorten wie Kuppenlagen, im Steilhang usw.
Organismengruppen als Zeiger für ökologische Einnischung
Pilze an Edelkastanie
 Zoombild vorhanden
Zoombild vorhanden
Abb. 11: Schwefelporling (Laetiporus sulphureus) an Buche, eine der Pilzarten mit starkem Einfluss auf die Edelkastanie (Foto: E. Segatz)
2016 erschien eine zusammenfassende Arbeit über »Pilze an [i]Castanea sativa[/i]« von J. A. Schmitt, der auch im Projekt »Die Edelkastanie am Oberrhein« verschiedene Standorte an der Haardt untersucht hat. Die folgenden Ergebnisse sind dieser verdienstvollen und in ihrer Gesamtdarstellung bisher einmaligen Arbeit entnommen. Ausgewertet wurden die eigenen Erhebungen von J. A. Schmitt sowie eine Vielzahl mykologischer Publikationen. Ziel war es, möglichst viele der mit [i]Castanea sativa[/i] assoziierten Pilztaxa sowohl der Mykorrhiza bildenden, als auch saprobiotischen und parasitischen Arten, Varietäten und Formen zu erfassen. Letztere kommen an Holz, Wurzeln, Rinde bzw. Borke, Blütenständen, Früchten, Fruchtschalen, Blättern und Streu vor. Nicht aufgenommen wurden von ihm Flechten und Flechten begleitende Pilzarten an der Borke lebender Edelkastanien.
Insgesamt wurden 805 Pilztaxa an [i]Castanea sativa[/i] dokumentiert. Alle Pilztaxa wurden hinsichtlich ihrer systematischen Zugehörigkeit und ihrer Ökologie aufgeschlüsselt. J. A. Schmitt beschreibt sieben neue Pilzarten bzw. -varietäten an [i]Castanea sativa[/i].
523 Pilzarten gehören zu saprobiotischen bzw. parasitischen Sippen. 41 % dieser Pilze sind Nichtblätterpilze, 23 % Schlauchpilze und 17 % Blätter- und Röhrenpilze.
Geht man von einer aus menschlicher, ökonomisch ausgerichteter Sicht »schädigenden« Pilzwirkung aus, können rund 82 % als harmlos eingestuft werden. Rund 14 % verursachen »geringe Schäden«.
Für »erhebliche Schäden« sind 13 Pilzarten, das sind rund 3 % aller dokumentierten Pilze, verantwortlich, darunter die bekannteren Arten [i]Armillaria mellea[/i] (Hallimasch), [i]Fistulina hepatica[/i] (Ochsenzunge) und [i]Fomes fomentarius[/i] (Echter Zunderschwamm).
»Starke Schäden«, die oft zum Absterben des Baumes führen, bewirken nur drei Arten (rund 0,6 % aller festgestellten Pilze), an erster Stelle [i]Cryphonectria parasitica[/i] (Edelkastanien-Rindenkrebs), dann [i]Phytophthora cambivora[/i], ein Erreger der Tintenkrankheit. Beide führen auch am Oberrhein zu flächenmäßig bedeutenden Ausfällen. Als weiterer Pilz dieser Gruppe wird [i]Laetiporus sulphureus[/i] (Schwefelporling) genannt (Abbildung 11).
Erstmals berichtet J. A. Schmitt auch über Mykorrhiza- Pilze, die mit der Edelkastanie in Symbiose leben. Da in der Literatur zu diesem Thema oft Daten aus Mischwäldern mit Edelkastanie vorliegen, können daraus nicht immer sichere Aussagen über die Mykorrhiza- Bindung von Pilzen an [i]Castanea sativa[/i] gemacht werden, weshalb in der Arbeit nur solche Pilztaxa aufgenommen wurden, die dezidiert [i]Castanea sativa[/i] als Mykorrhizabiont angeben.
Bei den 287 Mykorrhizabionten dominieren die Blätter- und Röhrenpilze mit 93 %. Diese sind nicht ausschließlich an [i]Castanea[/i] gebunden, sondern ganz überwiegend an Arten der [i]Fagales[/i], hier vor allem an nahe verwandte [i]Quercus[/i]-Arten, insbesondere die heimischen Arten [i]Quercus robur[/i] und [i]Quercus petraea[/i]. Mykorrhiza-Pilze sind von essenzieller Bedeutung für die Ernährung und die Wasserversorgung vieler Laubbäume und für die Stabilität entsprechender Waldökosysteme.
Flechten auf Edelkastanien
 Zoombild vorhanden
Zoombild vorhanden
Abb. 12: Physcia stellaris, eine Flechte an Edelkastanie (Foto: R. Cezanne/M. Eichler)
Insgesamt wurden 108 Taxa bestimmt, davon 99 eigentliche Flechten und neun lichenicole, also flechtenbewohnende Pilze, die von Flechtenkundlern mit aufgenommen werden. Die höchste an einer einzelnen Edelkastanie festgestellte Artenzahl betrug 55, im Mittel fanden sich 40 Flechtenarten pro Baum (Cezanne und Eichler 2012).
Nach Wuchsformen konnten 60 % den Krustenflechten, 33 % den Blattflechten (Abbildung 12) und 7 % den Strauchflechten zugeordnet werden. In der Krone wuchsen meist mehr Arten als am Stamm, 23 Arten ausschließlich in der Krone. 31 Arten lebten auf Totholz, davon jedoch nur zwei Arten ausschließlich.
Unter den Flechten fanden sich auch seltene Arten, zwölf Arten der Roten Liste für Deutschland und 27 Arten der Roten Liste für Rheinland-Pfalz. Fünf bisher in RLP unbekannte Arten wurden entdeckt, zwei Arten der Kategorie »ausgestorben oder verschollen« wiederentdeckt.
Dietrich und Bürgi-Meyer (2011) kommen für die »Chestenenweid« am Vierwaldstätter See im Kanton Luzern zu ähnlich reichhaltigen Befunden und äußern: »Die Edelkastanie stellt das wertvollste Substrat für baumbewohnende Flechten dar«.
Moose auf Edelkastanien
 Zoombild vorhanden
Zoombild vorhanden
Abb. 13: Rogers Goldhaarmoos. (Foto: M. Lüth)
Weitere elf Arten an Edelkastanie wurden bei einer erweiterten Suche auf feuchteren Standorten dokumentiert. Die höchste Artenzahl an einem Einzelbaum betrug 17, im Mittel zehn. Auch bei den Moosen fanden sich seltenere Arten, z. B. neun Arten der Roten Liste Rheinland-Pfalz und mit Rogers Goldhaarmoos [i](Orthotrichium rogeri)[/i] eine seltene, neue Art (Abbildung 13).
Mit Edelkastanie zusammenlebende Käfer
 Zoombild vorhanden
Zoombild vorhanden
Abb. 14: Cardiophorus gramineus, mit Ameisen zusammenlebend. (Foto: F. Köhler)
Zur Untersuchung wurde das Standardmethodenprogramm der rheinland-pfälzischen Naturwaldforschung aus Flugfallen, Leimringen, Totholzgesiebe und Klopstockfängen eingesetzt, lokal ergänzt um weitere Techniken wie Baumhöhleneklektoren, Lichtfallen, Flugköderfallen, Bodenfallen und Autokescher.
131 Proben enthielten 29.076 vollständig bis auf Artniveau bestimmte Käfer. An den vier Standorten wurden zwischen 278 und 571 Käferarten gefunden, insgesamt 1.002 Käferarten (Abbildungen 14, 15 und 16).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 15: Ampedus cardinalis (Urwaldreliktart und Pilzkäfer an Schwefelporling). (Foto: F. Köhler)
Lediglich rund 45 % der Käferarten sind an Waldbiotope gebunden, wobei ein auffällig hoher Anteil lichte Gehölzstrukturen bevorzugt (Köhler 2016) ein Hinweis auf die Behandlung alter Bestände unter dem Aspekt der Artenvielfalt.
200 Offenlandbewohner hängen mit der starken Auflichtung dreier Untersuchungsbestände zusammen. 329 Arten sind an Totholz gebunden, 103 Arten davon leben lignicol (Holzkäfer), 100 corticol (Rindenkäfer) oder succicol (Saftkäfer) und 55 Arten polyporicol (Pilzkäfer), 71 Arten sind xylodetriticol (Mulmkäfer) oder xylonidicol (Nestkäfer) (Köhler 2016).
Sowohl für die wenigen in der Literatur genannten als auch beobachteten 329 Xylobionten ergibt sich eine hohe Übereinstimmung mit der an Eiche [i](Quercus)[/i] bekannten Totholzkäferfauna. Im standardisierten Vergleich mit der Totholzkäferfauna rheinland-pfälzischer Naturwaldreservate erweisen sich die älteren Edelkastanienstandorte als ähnlich artenreich wie die international bedeutsamen Reservate im Bienwald (Köhler 2016).
Resumée
 Zoombild vorhanden
Zoombild vorhanden
Abb. 16: Hedobia regalis (Poch-, Klopf-, Bohr- oder Nagekäfer im Holz, wärmeliebend). (Foto: F. Köhler)
Auch die weiteren untersuchten Organismengruppen deuten in dieselbe Richtung. Das Inventar an Pilz-, Flechten- und Moosarten an Eiche ist weitgehend identisch mit dem der Edelkastanie. Insbesondere die große Anzahl an Mykorrhizapilzen belegt die Einnischung der Edelkastanie in die wärmeliebenden heimischen Waldökosysteme.
Zusammenfassung
Neben der Beschreibung von Aspekten der Beeinflussung des Standorts durch den Baum und von Besonderheiten der Fortpflanzungsökologie in Bezug auf die Insekten wird zudem versucht, die ökologischen Auswirkungen unterschiedlicher waldbaulicher Behandlungsmodelle grob zu skizzieren. Die Möglichkeiten, die die Edelkastanie für die Umsetzung eines Biotopholz-, Altholz- und Totholzprojekts bietet, werden anhand der dafür geschaffenen Elemente diskutiert.
Ausführlich werden Organismengruppen wie Pilze, Flechten und Moose sowie Totholzkäfer behandelt, die Hinweise auf das Einfügungsvermögen der Baumart in bestehende Waldökosysteme geben können.