


RSS-Feed der Bay. Landesanstalt für Wald und Forstwirtschaft abonnieren
So verpassen Sie keine Neuigkeiten mehr. Unser RSS-Feed "Nachrichten der Bayerischen Landesanstalt für Wald und Forstwirtschaft" informiert Sie kostenlos über unsere aktuellen Beiträge.
Nicole Burgdorf und Ludwig Straßer
Aktuelle pilzliche Erkrankungen bei Ahorn – LWF aktuell 124
Durch häufigere Dürre- und Hitzeperioden sowie eingeschleppte Arten nimmt die Gefährdung von Ahorn durch Pathogene zu.
Baumarten der Gattung Ahorn, insbesondere der weit verbreitete Bergahorn (Acer pseudoplatanus), galten bisher als robust und anbausicher. Steigende Temperaturen und mangelnde Niederschläge schwächen einerseits die Bäume, andererseits werden pilzliche Schaderreger begünstigt. Ein Auftreten verschiedener, teilweise neuer Krankheiten in den vergangenen Jahren ist die Folge.
Nach dem Trockensommer 2003 trat an Ahorn verstärkt die Verticillium-Welke in Erscheinung. Der eingeschleppte Erreger des Ahorn-Stammkrebses, dessen Schadpotenzial noch nicht abzuschätzen ist, wurde erstmals 2013 im Raum München nachgewiesen. Seit 2018 häufen sich in warmtrockenen Gebieten Schäden durch die Ahorn-Rußrindenkrankheit.
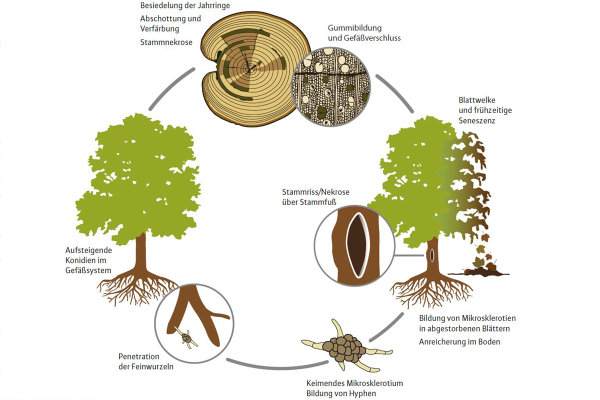 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Entwicklungszyklus des Pilzes Verticillium dahliae (Grafik: C. Hopf)
Die Bäume werden durch längere Dürreperioden und höhere Temperaturen geschwächt, wodurch physiologische Prozesse, die für Abwehrmechanismen gegenüber Schaderregern notwendig sind, nicht mehr aufrecht erhalten werden können. Zudem können sommerliche Klimaextreme, wie wir sie in den letzten Jahren erleben, zu irreversiblen Schädigungen führen, die die Vitalität insgesamt beeinträchtigen und eine Anfälligkeit erhöhen.
Das gemeinsame Auftreten einer klimabedingten Disposition der Bäume und eingeschleppter oder begünstigter Pflanzenpathogene führt derzeit bei der Gattung Ahorn, insbesondere auf ungeeigneten Standorten, zu einer Zunahme von Krankheiten.
Verticillium-Welke
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Endogener Stammriss und moosgrüne Verfärbungen mehrerer Jahrringe (Foto: N. Burgdorf)
In Südeuropa ist der Erreger insbesondere für große ökonomische Schäden im Olivenanbau verantwortlich. Der Pilz bildet langlebige Dauerorgane im Boden aus, die zehn Jahre und länger in Abwesenheit eines Wirts überleben können (Wilhelm 1955). Unter günstigen Bedingungen keimen diese Mikrosklerotien aus und können Pflanzen über die Feinwurzeln infizieren. Bei einer Infektion werden Blattchlorosen und Welke einzelner Kronenteile sichtbar, was im Extremfall zum Absterben der Pflanzen führt.
Die eher unspezifische Symptomatik ohne eine Ausbildung von Fruchtkörpern erschwert häufig eine eindeutige Diagnose. Bei der Infektion mit V. dahliae werden die Wasserleitbahnen des Baumes verstopft. Dadurch werden bei Ahorn im Querschnitt moosgrüne Verfärbungen einzelner Jahrringe sichtbar (Abbildung 1, 2), worüber der Erreger bei einer frischen Infektion isoliert und nachgewiesen werden kann.
Bei fortschreitender Krankheitsentwicklung können sich Stammrisse (Abbildung 3) und auch endogene Risse im Splintholz ausbilden, wobei oftmals mehrere Risse versetzt auftreten (Butin 2011). Als Ursache für die Entstehung der Stammrisse ist bei einer Infektion der zusätzliche Faktor Trockenstress anzusehen.
Bei intensiver Sommertrockenheit, die im Rahmen der prognostizierten klimatischen Veränderungen in Zukunft verstärkt auftreten kann, fällt es befallenen Wirtspflanzen schwerer, eine Infektion mit V. dahliae zu kompensieren. Neben dem akuten Krankheitsverlauf kann eine über Jahre verlaufende symptomlose chronische Phase auftreten.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Stammnekrose (Foto: N. Burgdorf,)
Bei jüngeren Pflanzen kann die Verticillium-Welke bei zusätzlichem Trockenstress zu einem schnellen Absterben führen. In Gewächshausstudien konnte ein additiver Effekt der Faktoren V. dahliae-Infektion und Trockenstress nachgewiesen werden. Bei der prognostizieren zunehmenden Sommertrockenheit muss damit gerechnet werden, dass bei einer bestehenden Verticillium- Infektion bei Ahornen vermehrt Stammschäden und Mortalität auftreten.
Eutypella-Stammkrebs
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: Ahorn-Stammkrebs mit Lagern der Fruchtkörper (Foto: V. Schlenker, LWF)
Im Herbst 2017 konnte Eutypella- Stammkrebs im Rahmen eines Monitorings der Abteilung Waldschutz (LWF) an Ahornen in Waldgebieten um München detektiert werden (Burgdorf et al. 2019). Im Sommer 2019 wurde ein weiterer Infektionsherd mit über 150 Bäumen in einem kleinen Laubmischwaldbestand im Oberallgäu entdeckt.
Beim Auftreten der Erkrankung scheint vornehmlich Bergahorn betroffen zu sein, doch konnten auch Nachweise an Spitz- und Feldahorn erbracht werden. Das Risiko der weiteren Ausbreitung von Ahorn-Stammkrebs wird aufgrund der Verbreitung der Ahornarten für Mitteleuropa als sehr hoch eingeschätzt (Ogris et al. 2006).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Ahorn-Stammkrebs im Querschnitt (Foto: N. Burgdorf)
Die Krebsbereiche erscheinen meist länglich elliptisch, abgeflacht und führen im Laufe der Jahre durch die Kallusbildung des Baumes zu einer Stammdeformation (Abbildungen 4, 5). Die Fruchtkörper des Pilzes werden erst nach fünf bis acht Jahren auf der Rinde gebildet (French 1969; Kliejunas 1971) und stellen eine dauerhafte Infektionsquelle dar.
Bei Niederschlägen und hoher Luftfeuchtigkeit werden die Sporen ausgeschleudert (French 1969; Johnson 1971). Aufgrund der Epidemiologie von E. parasitica sind somit Gebiete mit hohen Niederschlagsmengen und in Gewässernähe stärker gefährdet.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 6: Stammbruch nach fortgeschrittenem Befall von Ahorn-Stammkrebs (Foto: N. Burgdorf)
Bei der langjährigen Entwicklung des Krebses, insbesondere bei abnehmender Höhe am Schaft, erhöht sich daher die Stammbruchgefahr erheblich (Abbildung 6), was zu einer erhöhten Aufmerksamkeit in der Verkehrssicherungspflicht veranlasst.
Rußrindenkrankheit
 Zoombild vorhanden
Zoombild vorhanden
Abb. 7: Absterbende Kronen und starke Wasserreiserbildung (Foto: N. Burgdorf)
Seit dem verstärkten Auftreten seit 2005 (Metzler 2006; Kehr 2007; Robeck 2008) wurde das Pathogen in vielen Bundesländern detektiert. Das Auftreten der Erkrankung war bei den Erstnachweisen in Bayern im Sommer 2018 (Bork 2018) auf Waldbestände im warmtrockenen Unterfranken beschränkt. Seitdem ist eine weitere Ausbreitung und Intensivierung in Wäldern und in urbanen Bereichen zu beobachten.
Auffällig ist derzeitig das Auftreten in Bereichen von Waldinseln oder in städtischen Bereichen, wo Klimaextreme weniger abgeschwächt werden als in großflächigen, geschlossenen Wäldern. Die Symptomatik wurde vornehmlich bei Bergahorn in Rein- und Mischbeständen verschiedener Altersklassen festgestellt, aber auch Spitz-, Feld- und Silberahorn können erkranken.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 8: Bergahorn mit schuppenförmig abplatzender Rinde (Foto: N. Burgdorf, LWF)
Als erste Symptome sind vielfach Blattchlorosen und Welkesymptome zu beobachten, im weiteren Verlauf bilden sich Kronentotholz sowie verstärkt Wasserreiser im unteren Stammbereich aus (Abbildung 7). Der Pilz breitet sich im Holzkörper aus, was anhand von grün-bräunlichen Verfärbungen sichtbar wird (Abbildung 9).
Dringen diese bis in die Kambialbereiche vor, werden die schwarzbraunen Sporen massenhaft unter der Rinde gebildet. C. corticale ist zudem als latenter Endophyt beschrieben (Kelnarová et al. 2017), der wahrscheinlich lange Zeit in der Pflanze überdauern kann, bis es unter trocken-heißen Bedingungen zum Ausbruch der Erkrankung kommt.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 9: Verfärbungen im Kern- und Splintholz (Foto: N. Burgdorf)
Von den befallenen Bäumen mit ausgebildeten Sporenlagern war bereits ein Anteil von 90 Prozent abgestorben. Mit Sporenfängern konnte innerhalb und außerhalb zweier Untersuchungsbestände eine hohe Belastung mit den Konidiosporen nachgewiesen werden. Die Sporen von C. corticale werden vornehmlich im unteren Stammbereich gebildet und können auch noch nach der Fällung auftreten. Sind diese durch Wind und Niederschläge abgetragen, werden keine neuen Sporenlager mehr ausgebildet.
Als wichtigen Aspekt für die forstliche Praxis gilt es bei der Entnahme der Bäume zu beachten, dass die Sporen des Erregers bei intensivem Einatmen eine Gesundheitsgefahr darstellen können.
Klimaeinfluss
Dadurch können neue Pathogene begünstigt werden, aber auch das Schadpotenzial bekannter Forstpathogene reduziert werden. Ist ein Baum gestresst, steht weniger Energie für die Aufrechterhaltung der für die Krankheitsresistenz notwendigen physiologischen Prozesse zur Verfügung (Wargo & Harrington 1991). Krankheiten, die von einer schlechten Baumgesundheit profitieren, werden sich daher weiter durchsetzen (La Porta et al. 2008). Der Anbau von Bergahorn in trockeneren und wärmeren Gebieten Bayerns ist besonders in Zeiten des Klimawandels mit einem erhöhten Ausfallrisiko behaftet (Brosinger & Schmidt 2009) und in Zukunft als problematisch anzusehen.
Bislang ist wenig bekannt, wie der Faktor Dürre bzw. Wassermangel Einfluss auf Krankheitserreger ausübt und im Zusammenspiel zum Absterben von Bäumen führen kann (Sangüesa-Barreda et al. 2015). Viele Pilze benötigen für ihr Wachstum und zur Verbreitung Feuchtigkeit und könnten durch Dürreperioden weniger zu der Bildung von Myzel und Fruchtkörpern gelangen, wie beispielsweise der Erreger des Eschentriebsterbens in Folge des Trockensommers 2018. Andere pilzliche Schaderreger kommen erst unter trocken-heißen Bedingungen zum Zuge und könnten sich verstärkt etablieren und ausbreiten. Dies macht neben der Einschleppung von Arten durch den globalen Handel ein zunehmendes Auftreten neuer Krankheitsbilder wahrscheinlich.
Zusammenfassung
Die Schadintensität wird, wie bei der bereits seit längerem bekannten Verticillium- Welke, durch Hitze und Dürre begünstigt und kann in Zukunft den Anbau dieser Baumart auf bestimmten Standorten einschränken. Sollten die Pathogene bereits großflächig als Endophyten bei der Gattung Ahorn verbreitet sein, ist bei zukünftigen Klimaextremen mit einer zunehmenden Intensivierung des Schadgeschehens zu rechnen.
Eine weitere Bedrohung für den Bergahorn stellt wiederum in niederschlagsbegünstigten Gegenden der eingeschleppte Erreger des Eutypella- Stammkrebses an Ahorn dar. Die mangelnde Koevolution von Wirt und Parasit kann besonders unter disponierenden Bedingungen das Risiko einer Zunahme dieser neuartigen Erkrankung erhöhen. Um den zukünftigen waldbaulichen Anforderungen durch Klimawandel und Forstpathogene gerecht zu werden, sollte in Zukunft bei jeder Baumart intensiv auf die artspezifischen Standortsansprüche sowie bei Neubegründungen bzw. Pflanzungen auf die Eignung der Herkünfte geachtet werden.
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; Mcdowell, N. et al. (2010): A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259, S. 660–684
- climate change risks for forests. Forest Ecology and Management 259, S. 660–684 Bork, K. (2018): Rußrindenkrankheit an Ahorn – Erstfund in Bayern. AFZ–Der Wald 20, S. 34–35
- Brosinger, F., Schmidt, O. (2009): Beiträge zum Bergahorn. LWF Wissen 62, Freising, 75 S.
- Burgdorf, N.; Lemme, H.; Straßer, L. (2019): Aktueller Stand zur Verbreitung von Eutypella-Stammkrebs an Ahorn im Raum München. Jahrbuch der Baumpflege, S. 255–261
- 255–261 Butin, H. (2011): Krankheiten der Wald- und Parkbäume. Verlag Eugen Ulmer, Stuttgart, 318 S.
- Cech, T.L.; Schwanda, K.; Klosterhuber, M.; Straßer, L.; Kirisits, T. (2016): Eutypella canker of maple: first report from Germany and situation in Austria. Forest Pathology 46, S. 336-340
- Desprez-Loustau, M.L.; Marcais, B.; Nageleisen, L.M.; Piou, D.; Vannini, A. (2006): Interactive effects of drought and pathogens in forest trees. Annuals of Forest Science 63, S. 597–612
- Ellis, J.B.; Everhart, B.M. (1889): New species of hyphomycetous fungi. Journal of Mycology 5, S. 69
- French, W.J. (1969): Eutypella canker on Acer in New York. State Univ. College Forest., Syracuse Univ., Syracuse, Tech. Pub. 94, 56
- Gregory, P.H.; Waller, S. (1951): Cryptostroma corticale and sooty bark disease of sycamore (Acer pseudoplatanus). Mycological Research 34, S. 579-597
- Hiemstra, J.A.; Harris, D.C. (1998): A Compendium of Verticillium Wilts in Tree Species. Ponsen & Looijen, Wageningen, Niederlande, 80 S.
- Johnson, D.W. (1971): Biology, epidemiology, and control of Eutypella parasitica. Dissertation, University of Wisconsin-Madison, 133 S.
- Kehr, R. (2007): Neue Krankheiten an Platane, Linde und Ahorn. Jahrbuch der Baumpflege 2007, S. 144–156
- Kelnarová, I.; Černý, K.; Zahradník, D.; Koukol, O. (2017): Widespread latent infection of Cryptostroma corticale in asymptomatic Acer pseudoplatanus as a risk for urban plantations. Forest Pathology 47: e12344
- Kliejunas, J.T. (1971): Pathogenesis and developmental morphology of Eutypella parasitica in maple. Dissertation, University of Wisconsin-Madison, 117 S.
- La Porta, N.; Capretti, P.; Thomsen, I.M.; Kasanen, R.; Hietala, A.M.; Von Weissenberg, K. (2008): Forest pathogens with higher damage potential due to climate change in Europe. Canadian Journal of Plant Pathology 30, S. 177–195
- Metzler, B. (2006): Cryptostroma corticale an Bergahorn nach dem Trockenjahr 2003. Mitteilungen der Biologischen Bundesanstalt für Land- und Forstwirtschaft 400, S. 161–162
- Ogris, N.; Jurc, D.; Jurc, M. (2006): Spread risk of Eutypella canker of maple in Europe. EPPO Bulletin 36, S. 475–485
- Plate, H.-P.; Schneider, R. (1965): Ein Fall von asthmaartiger Allergie, verursacht von Cryptostroma corticale. Nachrichtenblatt des deutschen Pflanzenschutzdienstes 17, S. 100–101
- Robeck, P.; Heinrich, R.; Schumacher, J.; Feindt, R.; Kehr, R. (2008): Status der Rußrindenkrankheit des Ahorns in Deutschland. Jahrbuch der Baumpflege 2008, S. 238-245
- Sangüesa-Barreda, G.; Camarero, J. J.; Oliva, J.; Montes, F.; Gazol, A. (2015): Past logging, drought and pathogens interact and contribute to forest dieback. Agricultural and Forest Meteorology 208, S. 85–94
- Sinclair, W.A.; Lyon, H.H.; Johnson, W.T. (1987): Diseases of Trees and Shrubs. Comstock Publishing Associates, Cornell University Press, Ithaca, 576 S.
- Wargo, P.M.; Harrington, T.C. (1991): Host stress and susceptibility. In Armillaria root disease. Hrsg.: Shaw, C.G., Kile, G.A. USDA Forest Service in Agriculture 691, S. 88–101
- Wilhelm, S. (1955): Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology 45, S. 180–181