LWF Wissen 86
Aspekte zu Vermehrungsgut und Genetik der Buche
von Randolf Schirmer, Muhidin Šeho, Barbara Fussi, Andreas Ludwig und Michael Luckas

Abb. 1: Erntebestand »Höllbachschlag« im Nationalpark Bayerischer Wald (© M. Luckas)
Bei Buche als wichtigste heimische Laubbaumart kommt es entscheidend auf die Herkunft des verwendeten Vermehrungsguts an. Saatguterntebestände aus warm-trockenen Regionen werden an Bedeutung gewinnen. Bei dauerhaft ansteigenden Sommertemperaturen ist mit einem Nachlassen geeigneter Masten auf Grund von Trockenheit und Vitalitätsverlusten zu rechnen. Saatgutlagerung ist nur über kurze Zeiträume möglich. Neben Feldversuchen wird mittels ökologischer Nischenmodelle versucht, im Klimawandel besonders geeignete Herkünfte zu lokalisieren. Das Genom der Buche ist entschlüsselt, aber die Bestimmung adpativer Marker auf dem Genom, welche beispielsweise die Trockenheitsresistenz steuern, ist derzeit noch nicht möglich. Der Ausweisung von genetisch besonders wertvollen Beständen in Umsetzung des Generhaltungsprogramms kommt daher besondere Bedeutung zu.
Die Buche ist die dominierende Baumart der feucht-gemäßigten Klimazonen Europas. In Gebieten mit zunehmendem Kontinentalklima verliert sie wegen mangelnder Frostresistenz und ausgeprägter Spätfrostanfälligkeit ihre Konkurrenzkraft gegenüber Nadelholzarten. Auch bei verstärkt auftretender Trockenheit erreicht sie den Randbereich ihres natürlichen Vorkommens. In den letzten Trockensommern wurde ihr Vitalitätsverlust auf Grund ausbleibender Niederschläge besonders sichtbar. Obwohl ihr großes Verbreitungsgebiet auf eine hohe Klimaplastizität schließen lässt, gehört die Buche bei steigenden Temperaturen und nachlassenden Niederschlägen in den standörtlichen Randbereichen ihrer Verbreitung – wie zum Beispiel auf der Fränkischen Platte – zu den Verlierern im Klimawandel.
Saatgut und Vermehrung
Erntebestände
Gemäß Forstvermehrungsgutgesetz (FoVG) darf nur in amtlich zugelassenen und im Erntezulassungsregister eingetragenen Buchenbeständen Saatgut für den Vertrieb geerntet werden. Fachleute der Forstverwaltung wählen dafür Waldbestände aus, die die rechtlich geforderten Mindestkriterien erfüllen. Ein Alter von wenigstens 70 Jahren, eine Mindestbestandsgröße und vor allem Kriterien wie überdurchschnittliche Stammform, Wuchsleistung und Vitalität der Buchen sind entscheidend für die Zulassung. Durch Weitervererbung dieser Eigenschaften wird sichergestellt, dass Waldbesitzer gut veranlagte und leistungsfähige Pflanzen für ihren Wald erhalten.
Im Erntezulassungsregister (EZR) für Bayern, das am Bayerischen Amt für Waldgenetik (AWG) in Teisendorf geführt wird, sind 540 Saatguterntebestände mit einer baumartenbezogenen Fläche von 11.000 ha aufgeführt. Sie sind jeweils einem Gebiet mit annähernd einheitlichen ökologischen Bedingungen, dem sogenannten Herkunftsgebiet zugeordnet. Es handelt sich dabei um eine abgegrenzte Region, in der Saatguterntebestände ähnliche phänotypische und genetische Merkmale aufweisen. Gesundheit, Stabilität und Leistungsvermögen der Wälder hängen entscheidend von der Wahl der richtigen Herkunft ab.
Für die Rotbuche sind in Bayern 12 Herkunftsgebiete ausgewiesen. Zur Saatguternte zugelassene Buchenbestände sind in ganz Bayern zu finden, mit einem deutlichen Schwerpunkt im Spessart, der Fränkischen Platte und im Steigerwald. Ein besonders hochwertiger Buchensaatgutbestand in Hinblick auf Wuchs- und Formeigenschaften ist aber auch im Gebiet des Nationalparks Bayerischer Wald zu finden (Abb. 1). In Versuchen konnte eine Überlegenheit seiner Nachkommenschaften im Vergleich zu Vermehrungsgut aus Erntebeständen anderer Herkünfte nachgewiesen werden.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Männliche Blüte (links); weibliche Blüte (rechts) (© M. Luckas)
Frühestens im Alter von 30 Jahren beginnen Buchen, zeitgleich mit dem Blattaustrieb ab Ende April zu blühen. Der Witterungsverlauf im Vorjahr entscheidet darüber, ob aus einer Knospe ein Trieb oder eine Blüte wird. Trockenheit und hohe Temperaturen im Frühsommer können eine kräftige Blüte im Folgejahr verursachen. Buchen sind einhäusig – an einem Baum befinden sich männliche und weibliche Blüten. (Abb. 2). Sie stehen getrennt in unscheinbaren Blütenständen an jungen Trieben. Die männlichen Staubblüten hängen in Form büschelartiger Kätzchen an einem 3 bis 5 cm langen Stiel. Die weiblichen Blüten hingegen sitzen auf einem kurzen Stängel, meist zu zweit, aufrecht und geschützt von einer behaarten Umhüllung. Sie besitzen je einen Fruchtknoten mit drei Narben.
Samenreife und Ernte
Wie bei den meisten heimischen Waldbäumen erfolgt die Bestäubung auch bei der Buche durch Wind. Nach der Befruchtung der Samenanlagen entwickeln sich in einem stacheligen Fruchtbecher (Cupula) meist zwei dreikantige Nussfrüchte, die Bucheckern. Blüte und Fruktifikation finden allerdings nicht jedes Jahr statt, sondern in unterschiedlicher Intensität in nur unregelmäßigen Abständen. Im Zeitraum 2010/21 traten sechs Jahre ohne Erntemöglichkeiten auf (Abb. 3).
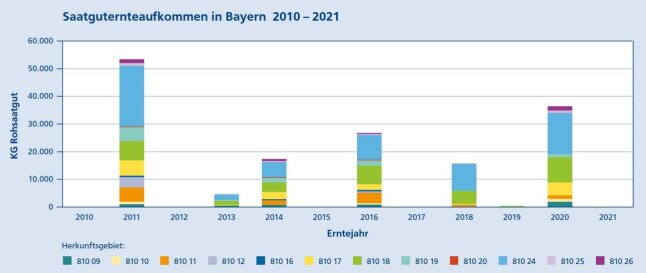
Abbildung 3: Entwicklung des Saatguternteaufkommens in Bayern im Zeitraum 2010 – 2021, aufgegliedert nach
Herkunftsgebieten Quelle: Bayer. Erntezulassungsregister
Der Reifevorgang der Früchte wird durch eine zunehmende Braunfärbung angezeigt und ist Mitte bis Ende September abgeschlossen. Mit fortschreitender Austrocknung springt der Fruchtbecher auf und lässt bis in den November hinein die glänzend braunen Bucheckern fallen. Nur nach einer üppigen Blüte der herrschenden und vorherrschenden Bäume und einer Fruchtreife ohne Spätfrost, Trockenheit, Sturm oder Hagelschlag ist die Voraussetzung für eine erfolgreiche Ernte gegeben. Alle 5 bis 8 Jahre können Buchenbestände sehr stark im Sinne einer Vollmast fruktifizieren. Dieser Begriff stammt aus dem Mittelalter, als Schweine in den Wald getrieben wurden, um sie dort mit Waldfrüchten zu mästen. Bucheckern bestehen zu 23 % aus Eiweiß und zu 50 % aus Fett. Menschen sollten keine größeren Mengen an Bucheckern verzehren, da die Samenhaut den giftigen Inhaltsstoff Fagin enthält, der zu Vergiftungen führen kann (Schütt 1992).
Je nach Intensität der Mast können von einem Baum 5 bis 25 kg Saatgut geerntet werden. Großkronige Solitärbäume können sogar bis zu 50 kg Bucheckern tragen. Bei Vollmasten liegen unter einem Baum bis zu 500 Samen/m² (Amann 1956).
Für die professionelle Saatgutbereitstellung werden von Forstsaatgutbetrieben Erntenetze unter den Samenbäumen ausgelegt, um die Bucheckern zu sammeln (Abb. 4).
Steckbrief Buchensamen| Fruchtbecher | Meist mit 2 Bucheckern |
| 1 kg Samen | ca. 5.000 Bucheckern |
| 1 kg Samen | ca. 800 – 15.00 Sämlinge |
| Tausendkorngewicht | ca. 190 – 220 g |
Wenn nach den ersten Herbststürmen im Oktober die Masse an Bucheckern gefallen ist, können die Netze zusammengezogen und das Buchensaatgut mittels spezieller Reinigungsmaschinen von Laub und sonstigen Verunreinigungen getrennt werden. Da im Frühherbst zuerst das Hohlkorn abfällt, sollten die Netze nicht zu früh ausgelegt werden.
In guten Erntejahren können in einem Bestand mehrere Tonnen Bucheckern gewonnen werden. Die rekordverdächtige Vollmast des Jahres 2011 führte bayernweit zu einem Ernteergebnis von etwa 53.000 kg gereinigten Saatguts. Aus Abbildung 3 wird deutlich, dass mehr als 50 % des in Bayern geernteten Buchensaatguts in den beiden Herkunftsgebieten Fränkische Alb (810 18) und Alpenvorland (810 24) gewonnen wird.
Saatgutbehandlung und Anzucht
Saatgutqualität und Lagerung
Die Qualität des Saatguts hängt u. a. von der Blühintensität und den klimatischen Bedingungen während der Fruchtreife ab. Eine Ernte ist nur lohnend, wenn 70 bis 80 % der Samen beim Aufschneiden ein weißes Endosperm aufweisen. Bei Vollmasten können bis zu 90 % der Bucheckern von hervorragender Qualität sein. Dagegen ist in Sprengmastjahren der Hohlkornanteil aufgrund von Selbstbestäubung häufig so hoch, dass von einer Beerntung abgesehen werden muss. Bei Spreng- und Halbmasten sind die wenigen Bucheckern zudem häufig von Schädlingen befallen und daher stark in ihrer Qualität beeinträchtigt.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Stratifikation von Bucheckern (links) und Aussaat (rechts) im BaySF Baumschulbetrieb Laufen (© A. Ludwig)
Buchensaatgut entwickelt durch Akkumulation verschiedener Hormone eine natürliche Keimhemmung (Bärtels 1996), wodurch die Keimung bis zum folgenden Frühjahr unterdrückt wird. Durch Stratifikation (Kalt-Nass-Behandlung) muss diese Keimhemmung spätestens vor der Aussaat überwunden werden. Hierzu wird das Saatgut bei einer Feuchte von 28 bis 32 % mindestens 100 Tage bei einer Temperatur von 3 bis 5 °C gelagert (Abb. 5). Danach kann das für die unmittelbar nachfolgende Frühjahrsaussaat vorgesehene Saatgut auf 20 bis 25 % Saatgutfeuchte getrocknet und bei – 3 °C zwischengelagert werden.
Um auch in Jahren ohne Erntemöglichkeiten ausreichend Saatgut zur Verfügung zu haben, können Bucheckern bei optimaler Ausgangsqualität und Aufbereitungsmethodik 3 bis 4 Jahre eingelagert werden. Niedriger Wassergehalt und geringe Lagertemperatur sind hierfür Voraussetzung. Das stratifizierte Saatgut wird daher auf 8 bis 10 % Feuchtegehalt getrocknet und in luftdicht verschlossenen PESäcken bei – 7 bis – 10 °C eingefroren. In Ausnahmefällen ist eine Lagerung bis zu 5 Jahren möglich (Suzka 1974).
Lagerversuche haben gezeigt, dass Saatgut aus Vollmasten länger gelagert werden kann als solches aus Halb und Sprengmasten (Schubert, o. A.) In einem Versuch wies 18 Jahre lang gelagertes Saatgut aus einer Vollmast noch 54 % Keimfähigkeit auf. Nicht unterschätzt werden darf die Erfahrung, dass länger gelagertes Saatgut im Tetrazoliumtest unter Laborbedingungen zwar noch über eine gute Keimfähigkeit verfügen kann, die Keimkraft zum Auflaufen unter Freilandbedingungen in der Baumschule jedoch stark nachgelassen hat. Ausfälle und geringes Sämlingswachstum sind die Folge. Saaten aus langfristiger Lagerung laufen im Saatbeet dann sehr lückig auf.
Buchensaatgut steht wegen der unregelmäßig auftretenden Masten und der begrenzten Lagerdauer nur in beschränktem Umfang zur Verfügung. Freisaaten können daher im Wald aufgrund des hohen Saatgutbedarfs von 200 – 300 kg/ha nur in Ausnahmefällen durchgeführt werden und beschränken sich meist auf Vollmastjahre.
In Baumschulen kann bei Aussaatmengen von 100 – 150 g/m2 mit 800 bis 1.500 verkaufsfertigen Sämlingen je Kilogramm Saatgut gerechnet werden (Krüssmann 1997). Als Regelsortiment werden ein bis zweijährige Pflanzen (1/0, 1/1 bzw. 2/0) in den Größenklassen 15/30, 30/50, 50/80 vom Waldbesitzer angefragt.
Herkunftsunterschiede entscheidend
Die Gattung Fagus wird anhand morphologischer Merkmale in die Arten Rotbuche (F. sylvatica) und in die in Kleinasien verbreitete Orientbuche (F. orientalis) abgegrenzt. Im Übergangsbereich beider Arten (Nordgriechenland, Südwestbulgarien, europäischer Teil der Türkei) tritt die Hybridform F. x moesiaca auf. In dieser Region kommt F. sylvatica vorwiegend in Berg lagen und F. orientalis in Tallagen und küstennahen Gebieten vor. Abbildung 6 zeigt die Herkunftsunterschiede anhand unterschiedlicher genetischer Muster.
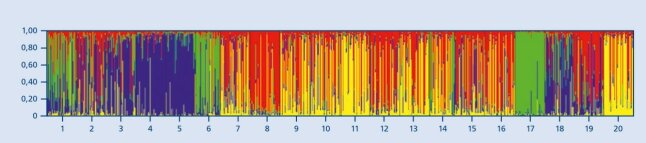
Abbildung 6: Genetische Unterschiede zwischen bulgarischen (1-6), deutschen (7-16), einer türkischen (17) einer
griechischen (18), einer italienischen (19) und einer schwedischen (20) Buchenpopulation (© LWF)
Buchen zeigen aufgrund ihres großen natürlichen Verbreitungsgebiets eine große Variabilität in ihren Leistungs und Qualitätsmerkmalen und in ihrer Toleranz gegenüber Spätfrost und Trockenheit. Bei der Pflanzung hat daher nicht nur die Herkunft, sondern ggf. sogar der jeweilige Saatguterntebestand einen großen Einfluss auf den Anbauerfolg. Die räumliche Nähe eines Saatguterntebestands zum Auspflanzungsort zeigte sich in Anbauversuchen als weniger wichtig für den Anbauerfolg als die Verwendung von Vermehrungsgut aus vergleichbaren Höhenlagen (Kleinschmit 2008).
Bei den genetisch beeinflussten Eigenschaften ist zwischen herkunftsbezogenen und qualitätsbezogenen Merkmalen zu unterscheiden.
Wichtigste herkunftsgesteuerte Merkmale sind Austriebszeitpunkt und -geschwindigkeit sowie der Vegetationsabschluss. Sie sind Ausdruck der regionalen Anpassung einer Herkunft. Werden Hochlagenherkünfte im Tiefland ausgepflanzt, treiben sie schneller aus als dort wachsende Herkünfte und sind somit stärker spätfrostgefährdet. Als Reaktion auf den Klimawandel ist daher ausschließlich ein begrenztes Verbringen von Tieflagenherkünften in etwas höhere Lagen ratsam.
Tieflagenherkünfte benötigen eine hohe Wärmesumme und zeigen somit im Frühjahr einen verzögerten Austrieb, der sie vor Spätfrösten schützt. Tieflagenherkünfte und südliche Herkünfte können die Vegetationszeit besser ausnutzen. In Versuchen korrelierte das Wachstum positiv mit der Zeit des Vegetationsabschlusses, aber nicht mit dem Vegetationsbeginn. Hochlagenprovenienzen beziehungsweise Herkünfte aus dem nördlichen Randbereich der Verbreitung erwiesen sich als wuchsschwächer, sind aber winterfrostresistenter (Rohmeder 1959). Sie treiben, wie Herkünfte aus südlicheren Lagen, schneller aus, wenn sie in Tieflagen oder nach Norden verbracht werden und sind somit stark spätfrostgefährdet. Die Herausforderung im Klimawandel wird sein, trockenheits und hitzetolerante Herkünfte zu finden, die jedoch trotzdem noch eine ausreichende Frosthärte aufweisen.
In Trockenstressuntersuchungen verschiedener Buchenherkünfte unter Laborbedingungen wurden erhebliche Unterschiede in der Reaktion einzelner Herkünfte beobachtet. Zentralpolnische Herkünfte litten weniger unter Trockenheit als Herkünfte aus dem ostdeutschen Tiefland und Westpolen (Czajkowski 2006).
Provenienzen aus wintermildem Küstenklima wachsen in kalten, subkontinentalen Regionen langsamer. Neben den herkunftsbezogenen Eigenschaften sind bei der Buche qualitätsbezogene Merkmale wie Drehwuchs, Zwieselbildung und Geradschaftigkeit in hohem Maß genetisch beeinflusst. Herkünfte aus höheren Lagen erbrachten in Anbauversuchen tendenziell bessere Schaftformen als Tieflagenherkünfte . Bei den Varietäten der Buche sind die rötlich gefärbten Blätter der Blutbuche (Abb. 7) sowie die herabhängenden Äste der Trauerbuche Beispiele für spontan auftretende Mutationen.
Anpassung der Verwendungsempfehlungen im Klimawandel
Die Trockenheit der letzten Jahre führte bei Buchenbeständen zu Vitalitätsverlusten und Absterbeprozessen, obwohl die Buche in Bayern vielerorts klimatische Optimalbedingungen vorfindet. Die Ergebnisse eines internationalen Herkunftsversuchs zeigen auf süddeutschen Versuchsflächen deutliche Unterschiede zwischen untersuchten Herkünften (Šeho et al. 2022). Bestimmte Herkünfte zeigen als »Generalisten« sowohl unter kalt-trockenen (Fichtelberg/Fichtelgebirge) als auch unter warm-trockenen (Kaiserstuhl/Freiburg) Klimabedingungen überdurchschnittliche Wuchsleistungen. Im Gegensatz dazu weisen »Spezialisten« nur unter kalttrockenen, kontinentalen Bedingungen eine höhere Überlebenswahrscheinlichkeit und ein erhöhtes Wachstum auf.
Neben Feldversuchen stellen nach Herkunftsregionen aufgesplittete Nischenmodelle eine neue Möglichkeit der Bewertung von Herkünften dar (Mellert et al. 2021a, b). Diese Vorgehensweise ermöglicht eine Aufgliederung der Herkunftsregionen nach dem dort herrschenden klimatischen Selektionsdruck. Dadurch kann die potenzielle Umweltanpassung auf der Ebene des Ökotyps bewertet werden.
Die Gliederung der europäischen Buchenherkunftsregionen erfolgt anhand der natürlichen Vegetation in 30 Einheiten (Abb. 8). Für diese Einheiten wird mit Methoden der Genetik, Baumphysiologie und Resilienzforschung untersucht, ob es unter heimischen Herkünften besonders klimatolerante, d. h. angepasste und anpassungsfähige lokale Herkünfte gibt, für die in der Praxis auch verfügbare Saatguterntebestände vorhanden sind.
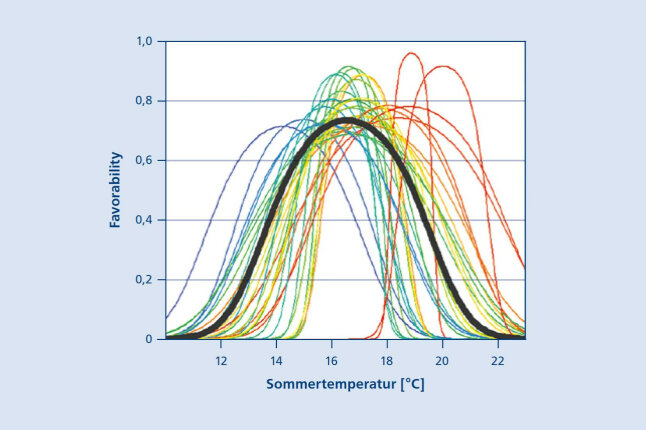
Abbildung 8: Nischenmodell für Buche (Mellert et al. 2021): Standortseignung (Favorability) unterschiedlicher Buchenherkünfte in Abhängigkeit von der Sommermitteltemperatur (© LWF)
Die Ergebnisse von forstgenetischen Feldversuchen und Modellierungen zeigen, dass ausgewählte Buchenherkünfte aus anderen Regionen Europas geeignet scheinen, um heimische Wälder mit an den Klimawandel angepasstem Saat und Pflanzgut anzureichern. Die aktualisierten Verwendungsempfehlungen sehen daher für besonders vom Klimawandel betroffene Herkunftsgebiete wie beispielsweise dem Oberrheingraben die Möglichkeit vor, im Rahmen von Praxisanbauversuchen Saatgut aus wärmeren Regionen (Nordostfrankreich, Burgund) testweise zu verwenden.
Genom entschlüsselt
Das Genom der Buche als wichtigste heimische Laubbaumart wurde bereits 2018 entschlüsselt (Mishra et al. 2018). Die vollständige DNA-Sequenz ist 542 Megabasen lang und besteht aus 542.000.000 Einzelbausteinen, die auf 12 Chromosomen verteilt sind. Insgesamt wurden 130.000 Rotbuchen-Gene identifiziert.
Neben dem Erbgut im Zellkern befinden sich auch in Chloroplasten und Mitochondrien weitere Erbinformation. Die Sequenz dieses mitochondrialen Genoms wurde ebenfalls entschlüsselt (Mader et al. 2020). Zu einer einzigen DNA-Sequenz zusammengesetzt, ergibt es eine Länge von 504.715 Basenpaaren.
Insgesamt wurden 58 Gene gefunden. Innerhalb dieser Gene konnten vier für Fagus spezifische Genmarker (sog. SNPs, Single Nucleotide Polymorphisms) entwickelt werden. Dieses Markerset eignet sich zur Identifizierung der Gattung Fagus in DNA-Proben aus Baumgeweben oder Holzprodukten.
Gene sind die Grundlage des Erscheinungsbildes eines Baumes. Je vielfältiger seine genetische Ausstattung, desto reaktionsfähiger ist er angesichts sich ändernder Umweltfaktoren. Auf den Bestand bezogen sagt die genetische Vielfalt etwas über die Unterschiede zwischen den Individuen aus und ist vergleichbar mit einem Werkzeugkoffer. Je mehr Werkzeuge zur Verfügung stehen, desto besser kann sich ein Bestand an neue Umweltbedingungen anpassen.
Genetische Muster und Rückwanderungswege
Die Buchenvorkommen in Europa lassen sich auf Grundlage der nacheiszeitlichen Rückwanderung in drei Hauptgenpools unterteilen. Es wird vermutet, dass die bayerischen Herkünfte vor allem aus dem nördlichen Balkan stammen. (Magri et al. 2006, Postolache et al. 2021).
Aufgrund der Ausbreitung des Pollens durch Wind und der großflächigen Vorkommen ist die genetische Differenzierung in Deutschland zwischen Regionen sehr gering. Innerhalb der Bestände ist jedoch eine hohe genetische Diversität festzustellen (Postolache et al. 2021). Daher ist bei der Pflanzung von Buchenbeständen die Verwendung von Saatgut aus möglichst vielen verschiedenen Beständen ratsam. Um die genetische Vielfalt im Saatgut zu erhöhen, eignet sich auch die gezielte Auswahl von vielen Plusbäumen und deren Zusammenstellung in möglichst großen Samenplantagen.
Anpassungsrelevante Marker – Studien zu Trockenstresstoleranz und Phänologie
Die Lokalisierung anpassungsrelevanter Gene, die für hohe Trockenstressresistenz verantwortlich sind, wird vor dem Hintergrund des Klimawandels immer wichtiger. Mittels dieser adaptiven Marker soll es in Zukunft gelingen, Saatgut von Buchenbeständen zu empfehlen, das mit ausgeprägter Sommertrockenheit besser zurechtkommt. Über Genexpressions- oder Assoziationsstudien werden phänotypische Merkmale und Reaktionen auf Stressbedingungen mit genetischen Markern in Verbindung gebracht. Unterschiede in einem Trockenstressgen wurden zwischen Beständen von trockenen und feuchten Standorten in Bulgarien festgestellt (Fussi et al. 2014). Nur wenn bestimmte Gene und Genkombinationen gefunden werden, die ursächlich für die Trockenstresstoleranz verantwortlich sind, kann eine Auslesezüchtung in Kombination mit Feldversuchen zu praxistauglichen Ergebnissen führen.
Die großräumige genetische Struktur der Buche zeigt, dass Bestände in hohem Maß über den Genfluss miteinander verbunden sind. Die Kombination aus Genfluss, der die Diversität innerhalb der Population aufrechterhält, und Selektion, die die adaptive Differenzierung zwischen Populationen bedingt, sichert die Grundlage für eine hohe Anpassungsfähigkeit (Postolache et al. 2021).
Zusätzlich können Umwelteinflüsse Gene aktivieren oder deaktivieren. Sobald diese Genregulation vererbt wird, spricht man von Epigenetik. Ein Gen für eine bestimmte Ausprägung eines Merkmals muss aber vorhanden sein. Es kommt auf das Zusammenspiel der Gene und deren Aktivitätszustand an, um in geeigneter Weise auf Umwelteinflüsse reagieren zu können. Viele der in einem Einzelbaum aufgebauten spezifischen Zustände der Gene werden im Zuge der Vererbung wieder gelöscht. Der Einfluss vererbter Muster der Genregulationen auf den Phänotyp und dessen Reaktionsfähigkeit muss erst geklärt werden. Bei der Buche ist über diese Weitergabe der Stresserinnerung noch wenig bekannt. Es gibt jedoch Untersuchungen, die darauf hinweisen, dass z. B. bestimmte Blattmerkmale (Spaltöffnungsgröße, Spaltöffnungsdichte, Blattmasse je Fläche) epigenetisch beeinflusst sein könnten (Petrik et al. 2022). So könnte ein Trockensommer im Folgejahr die Ausbildung von kleineren Spaltöffnungen, einer höheren Spaltöffnungsdichte und dickeren Blättern verursachen.
Generhaltung und genetisches Monitoring
Die Erhaltung forstlicher Genressourcen bei Buche erfolgt in Umsetzung des »Konzepts zur Erhaltung und nachhaltigen Nutzung forstlicher Genressourcen in Bayern« (ASP, 2015). Ziel ist die Sicherstellung der langfristigen Weitergabe der Erbinformationen an die Folgegenerationen. Die aktuellen Umweltveränderungen vollziehen sich so schnell, dass es zum Absterben von Buchenbeständen kommt und eine natürliche Anpassung nicht mehr möglich ist. Daher muss die Förderung der Biodiversität vorrangig in durch Trockenheit gefährdeten Regionen verstärkt werden, da sie die Grundlage der Anpassungsfähigkeit darstellt.
Die Ergebnisse zeigen eine hohe genetische Vielfalt innerhalb der untersuchten Bestände. Sie sind Grundlage der Empfehlungen für Maßnahmen zur Sicherung des Genpools in den beiden südbayerischen Generhaltungszonen, die 45 % der bayerischen Waldfläche repräsentieren. In den kommenden Jahren werden weitere Bestände in den drei nordbayerischen Generhaltungszonen ausgewählt und untersucht. Neben der Erhaltung der Buche in Wirtschaftswäldern sollen zusätzlich besonders schützenswerte Einzelvorkommen auf Sonderstandorten und in Naturwaldreservaten als Generhaltungsbestände ausgewiesen werden. Vorhandene Bestände sollen erhalten und besonders wertvolle Vorkommen repräsentativ beerntet und auf geeigneten Standorten neu etabliert werden.
Die Sicherung von Waldbeständen unter sich schnell ändernden Umweltbedingungen ist ein wichtiger Aspekt nachhaltiger Waldbewirtschaftung. Grundlage hierfür ist eine Langzeitbeobachtung der genetischen Variation. In wiederholten Inventuren wird dabei bewertet, ob ein Waldbestand auf lange Sicht überleben, sich fortpflanzen und fortbestehen kann (Fussi et al. 2016). Das AWG hat hierfür ein Monitoringkonzept entwickelt, auf dessen Grundlage schädliche Änderungen der genetischen Anpassungsfähigkeit der Wälder frühzeitig erkannt werden. Grundlage hierfür sind Dauerbeobachtungsflächen unter verschiedene Umweltbedingungen im Kranzberger Forst/Freising sowie in einem Bergwaldbestand bei Adlgaß/Inzell (Abb. 9). Neben regelmäßigen genetischen Untersuchungen werden phänologische Beobachtungen (Austriebsverlauf, Blüh- und Fruktifikationsintensität) sowie Vitalitätseinschätzungen an ausgewählten Bäumen durchgeführt. Zusätzlich werden auf beiden Flächen Wetterdaten aufgezeichnet. Dadurch werden z. B. Spätfröste, die sich negativ auf das Reproduktionsgeschehen (z. B. Absterben der Blüten) auswirken, dokumentiert.
In beiden Dauerbeobachtungsbeständen herrscht ein intakter Genfluss. Die genetische Vielfalt der Elternbäume wird vollständig auf die Naturverjüngung übertragen (Abb. 10).
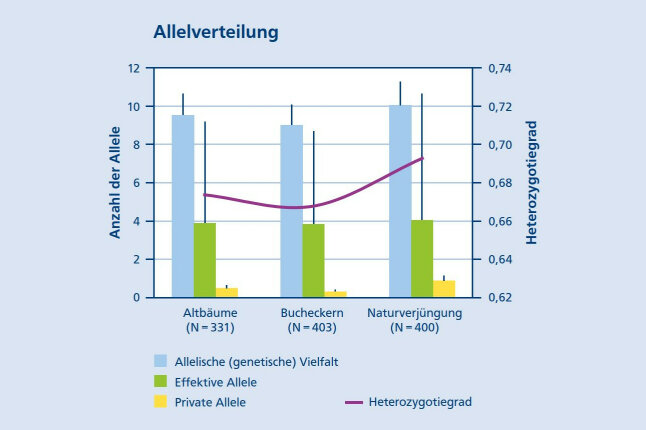
Abbildung 10: Genetische Vielfaltswerte für drei Generationen der Dauerbeobachtungsfläche Adlgaß (© LWF)
Auf der Buchenfläche Adlgaß wird eine erhöhte genetische Vielfalt bei der Naturverjüngung beobachtet, da die Pflanzen aus mehreren Samenjahren stammen. Durch das forstgenetische Monitoring können zeitliche und räumliche Veränderungen genetischer Parameter in den beiden Beständen frühzeitig aufgezeigt und die Anpassungsfähigkeit von Waldbeständen wissenschaftlich beurteilt werden.
Oberstes Ziel ist die langfristige Erhaltung der genetischen Information wertvoller Buchenbestände am Ort ihres Vorkommens. Die Bestände werden in sog. Generhaltungszonen nach ökologischen und genetischen Kriterien vorausgewählt. Derzeit läuft dieser Prozess in potenziellen Erhaltungsbeständen südlich der Donau bis hin zum Alpenbereich. Genetische Analysen von zehn Beständen ermöglichen einen Überblick über die genetische Variabilität sowie die räumliche Verteilung der genetischen Muster.
Summary
For beech as the most important native broadleaf tree species, the origin of the propagation material used is crucial. Seed stands from warm-dry regions will gain in importance. With permanently rising summer temperatures, a decline of suitable masts due to drought and loss of vitality is to be expected. Seed storage is only possible for short periods. In addition to field trials, eco-logical niche models are being used to try to localise par-ticularly suitable provenances under climate change. The genome of beech has been decoded, but the determination of adaptive markers on the genome, which control drought resistance, for example, is not yet possible. The designation of genetically particularly valuable stands in implementation of the gene conservation programme is therefore of particular importance.
Literatur
- Amann, G. (1956): Bäume und Sträucher des Waldes. Neumann- Neudamm, Melsungen.
- Bayerisches Amt für forstliche Saat und Pflanzenzucht (ASP) (2015): Konzept zur Erhaltung und nachhaltigen Nutzung forstlicher Genressourcen in Bayern. Kurzversion 23 S.
- Bärtels, A. (1996): Gehölzvermehrung. Ulmer. Stuttgart
- Czajkowski, T. und Bolte, A. (2006): Unterschiedliche Reaktion deutscher und polnischer Herkünfte der Buche auf Trockenheit. Allg. Forst- u. J.-Ztg. 177, S. 30-40
- Fussi, B., Cremer, E., Konnert, M. (2014): Auf der Suche nach anpassungsrelevanten Genen bei Waldbäumen. LWF Wissen 74, 27-31.
- Fussi, B., Westergren, M., Aravanopoulos, F., Baier, R., Kavaliauskas, D., Finzgar, D., Alizoti, P., Bozic, G., Avramidou, E., Konnert, M., Kraigher, H. (2016): Forest genetic monitoring: an overview of concepts and definitions. Environ Monit Assess 188:493; DOI 10.1007/s10661-016-5489-7.
- Kleinschmit et al (2008): Schutz durch Nutzung forstlicher Genressourcen der Buche in Nordwestdeutschland in: Nordwestdeutsche Forstliche Versuchsanstalt (Hrsg.): Ergebnisse angewandter Forschung zur Buche.
- Krüssmann, G. (1997): Die Baumschule Blackwell Wissenschaftsverlag, 6. Auflage.
- Mader, M., Schroeder, H., Schott, TH., Schöning-Stierand, K., Montalvao, A., Liesebach, H., Liesebach, M., Fussi, B., Kersten, B. (2020): Mitochondrial Genome of Fagus sylvatica L. as a Source for Taxonomic Marker Development in the Fagales. Plants, 9, 1274; doi:10.3390/plants9101274.
- Mellert, K.-H., Janßen, A., Šeho, M. (2021a): Anpassung an Klima und Boden bestimmt die Eignung von Herkünften. LWF aktuell 4, 43-45.
- Mellert, K.-H., Janßen, A., Šeho, M. (2021b): Wo finden wir Alternativherkünfte der Buche für den Klimawandel? AFZ-DerWald 24, 16 -20.
- Mishra B, Ulaszewski B, Meger J, Aury J, Bodénès C, Lesur-Kupin I, Pfenninger M, Silva CD, Gupta DK, Guichoux E, Heer K, Lalanne C, Labadie K, Opgenoorth L, Ploch S, Provost GL, Salse J, Scotti I, Wötzel S, Plomion C, Burczyk J, Thines M. (2018): A chromosome-level genome assembly of the European Beech (Fagus sylvatica) reveals anomalies for organelle DNA integration, repeat content and distribution of SNPs. doi:10.1101/2021.03.22.436437. PPR: PPR300961.
- Petrik, P., Petek‐Petrik, A., Kurjak, D., Mukarram, M., Klein, T., Gömöry, D., Střelcová, K., Frýdl, J., Konôpková, A. (2022): Interannual adjustments in stomatal and leaf morphological traits of European beech (Fagus sylvatica L.) demonstrate its climate change acclimation potential. Plant Biology.
- Postolache, D., Oddou-Muratorio, S., Vajana, E., Bagnoli, F., Guichoux, E., Hampe, A., Le Provost, G., Lesur, I., Popescu, F., Scotti, I., Piotti, A., Vendramin, G.G. (2021): Genetic signatures of divergent selection in European beech (Fagus sylvatica L.) are associated with the variation in temperature and precipitation across its distribution range. Mol Ecol 30:5029–5047. https://doi.org/10.1111/mec.16115
- Rohmeder, E. Schönbach, H., (1959): Genetik und Züchtung der Waldbäume. Parey, Hamburg, Berlin.
- Schubert, J. (o.A.): Lagerung und Vorbehandlung von Saatgut wichtiger Baum- und Straucharten; Landesanstalt f. Ökologie, Bodenordnung und Forsten, NRW, Eberswalde-Finow 183 S.
- Schütt, P.; Schuck H.J.; Stimm, B. (Hrsg.) (1992): Lexikon der Forstbotanik, ecomed, Landsberg/Lech.
- Šeho, M., Sommer, C., Kohnle, U. (2022): Der internationale Buchenherkunftsversuch von 1996/1998: Wachstums- und qualitätsrelevante Merkmale unter unterschiedlichen Standorts- und Klimabedingungen in Süddeutschland. Allg. Forst und J.-Ztg. 191 Jg. 11/12, 243-261.
- Suzka, B. (1974): Storage of beech for up to 5 winters, Arboretum Kornickie, 105-128.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren



 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
