


Randolf Schirmer und Eva Cremer
Aspekte zu Vermehrungsgut und Genetik der Robinie – LWF Wissen 84
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Robinienblüte (Foto: D. Schneck)
Die Robinie wurde in Frankreich bereits 1601 wegen ihrer auffallenden Blüte aus Nordamerika eingeführt. Schwerpunkt ihres Anbaus lag zunächst in Parks. Forstlich wurde ihr Anbau in Deutschland vor allem auf den armen Sandböden in Brandenburg gefördert, da sie mit Trockenheit zurechtkommt und die Böden als stickstoffbindende Art verbessert.
Heute sind vor allem ihre Eigenschaften als ökologisch interessanter Baum von Interesse. Neben landschaftsästhetischen Aspekten spielt sie als Bienenweide, zur Befestigung erosionsgefährdeter Böden und als kurzumtriebstaugliche Baumart eine Rolle.
Im Rahmen der Suche nach trockenverträglichen Baumarten im Klimawandel wird sie für forstliche Anbauten zunehmend an Bedeutung gewinnen. Vor diesem Hintergrund ist es wünschenswert, wenn das Vermehrungsgut eine hohe genetische Diversität aufweist.
In Bayern gibt es nur zwei nach Forstvermehrungsgutrecht zugelassene Erntebestände. Diese wurden jedoch noch nicht beerntet, da die Erntemöglichkeiten in Ostdeutschland wesentlich besser sind.
Heute sind vor allem ihre Eigenschaften als ökologisch interessanter Baum von Interesse. Neben landschaftsästhetischen Aspekten spielt sie als Bienenweide, zur Befestigung erosionsgefährdeter Böden und als kurzumtriebstaugliche Baumart eine Rolle.
Im Rahmen der Suche nach trockenverträglichen Baumarten im Klimawandel wird sie für forstliche Anbauten zunehmend an Bedeutung gewinnen. Vor diesem Hintergrund ist es wünschenswert, wenn das Vermehrungsgut eine hohe genetische Diversität aufweist.
In Bayern gibt es nur zwei nach Forstvermehrungsgutrecht zugelassene Erntebestände. Diese wurden jedoch noch nicht beerntet, da die Erntemöglichkeiten in Ostdeutschland wesentlich besser sind.
Blüte und Fruktifikation
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Blühender Robinienbestand (Foto: D. Schneck)
Robinien blühen alle ein bis zwei Jahre intensiv. Ihre Blüten erscheinen erst nach Laubausbruch Anfang Juni. Die Blütedauer mit 5 – 6 Tagen ist nur kurz. Der Blütenstand besteht aus einer 10 bis 25 Einzelblüten umfassenden Traube. Ihre leuchtend weißen Farben und der besondere Duft sind eine auffallende Besonderheit dieser Baumart und Grundlage für die Bestäubung durch Bienen.
Ein Robinienbestand von einem Hektar liefert die Grundlage für jährlich etwa 200 kg Honig (Mann 1989) (Abbildungen 1 und 2).
Ein Robinienbestand von einem Hektar liefert die Grundlage für jährlich etwa 200 kg Honig (Mann 1989) (Abbildungen 1 und 2).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Zugelassener Robinienbestand (Foto: V. Schneck)
Bis zum Herbst entwickelt sich aus dem Blütenstand eine maximal 10 cm lange und 2 cm breite, braune Hülse. Sie enthält 4 bis 10 nierenförmige Samen, die erst im Laufe des Winters entlassen werden. Die Samen sind olivgrün bis braun und oftmals marmoriert. Gelbe oder rote Samen sind unreif oder nicht keimfähig.
Ein Baum mittlerer Größe trägt 6 – 12 kg Saatgut. 100 kg reife Hülsen enthalten 15 – 33 kg Samen (Schütt 2008).Im Freistand fruktifizieren Robinien bereits ab einem Alter von zehn bis zwanzig Jahren, im Waldbestand erst ab dem Alter von dreißig Jahren.
Auf natürlichem Weg vermehren sich Robinien meist über Wurzelbrut, die aus Adventivknospen junger Horizontalwurzeln erwachsen. Sie kommen in einem Radius von bis zu 35 m um den Mutterbaum vor. Nach Fällung des Mutterbaums bzw. nach Verletzungen der Wurzeln entsteht Wurzelbrut besonders intensiv. Die Robinie gilt daher als invasive Baumart.
Ein Baum mittlerer Größe trägt 6 – 12 kg Saatgut. 100 kg reife Hülsen enthalten 15 – 33 kg Samen (Schütt 2008).Im Freistand fruktifizieren Robinien bereits ab einem Alter von zehn bis zwanzig Jahren, im Waldbestand erst ab dem Alter von dreißig Jahren.
Auf natürlichem Weg vermehren sich Robinien meist über Wurzelbrut, die aus Adventivknospen junger Horizontalwurzeln erwachsen. Sie kommen in einem Radius von bis zu 35 m um den Mutterbaum vor. Nach Fällung des Mutterbaums bzw. nach Verletzungen der Wurzeln entsteht Wurzelbrut besonders intensiv. Die Robinie gilt daher als invasive Baumart.
Zulassung, Beerntung und Ernteaufkommen
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: Zugelassener Robinienbestand (Foto: V. Schneck)
Die Robinie unterliegt seit 2003 dem Forstvermehrungsgutgesetz. Die Erzeugung und Vermarktung von Saat- und Pflanzgut ist daher nur aus zugelassenen Erntebeständen oder Samenplantagen erlaubt. Deutschland ist in die Herkunftsgebiete »Norddeutsches Tiefland (819 01)« und »Übriges Bundesgebiet (819 02)« unterteilt. Aktuell sind 37 Bestände mit einer Fläche von 112 ha in ganz Deutschland zugelassen. Mehr als 90 % dieser Bestände liegen in Brandenburg (Abbildung 3 und 4).
Die Aufgliederung in nur zwei Herkunftsgebiete ist ausreichend, da bei dieser nordamerikanischen Baum- art noch keine weitreichenden Anpassungsvorgänge unterstellt werden und sich die waldbauliche Bedeutung derzeit auf nur wenige Regionen beschränkt. Im bayerischen Staatswald wird die Anbaufläche auf 70 – 80 ha geschätzt (Biermayer 1999).
Als Mindestanforderung für die Zulassung als Erntebestand in der Kategorie »Ausgewählt« ist ein Alter von dreißig Jahren notwendig. Es können nur Bestände mit mindestens zwanzig Bäumen zugelassen werden. Diese sollten sich durch überdurchschnittliche Wipfelschäftigkeit, Feinastigkeit und einen geringen Anteil an Tiefzwieseln auszeichnen.
Die Aufgliederung in nur zwei Herkunftsgebiete ist ausreichend, da bei dieser nordamerikanischen Baum- art noch keine weitreichenden Anpassungsvorgänge unterstellt werden und sich die waldbauliche Bedeutung derzeit auf nur wenige Regionen beschränkt. Im bayerischen Staatswald wird die Anbaufläche auf 70 – 80 ha geschätzt (Biermayer 1999).
Als Mindestanforderung für die Zulassung als Erntebestand in der Kategorie »Ausgewählt« ist ein Alter von dreißig Jahren notwendig. Es können nur Bestände mit mindestens zwanzig Bäumen zugelassen werden. Diese sollten sich durch überdurchschnittliche Wipfelschäftigkeit, Feinastigkeit und einen geringen Anteil an Tiefzwieseln auszeichnen.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Samenplantage von Robinie in Sachsen-Anhalt (Foto: D. Schneck)
Das überragende Merkmal für eine Zulassung ist jedoch die Geradschaftigkeit, da viele Robinienbestände unschürige Stammformen aufweisen. Auffallend ist, dass vor allem stark blühende Individuen oft durch einen besonders schlängelnden Stamm gekennzeichnet sind.
In Bayern gibt es nur zwei Erntebestände im Raum Neustadt/Aisch und östlich von Bamberg. Sie wurden jedoch noch nie beerntet.
Bei einer Ernte muss auf jeden Fall von zehn Bäumen eines Bestands Saatgut eingesammelt werden, damit eine ausreichende genetische Vielfalt sichergestellt wird. Die Ernte erfolgt im Zeitraum November bis Februar durch Schütteln auf Netze bzw. durch Pflücken der Hülsen am stehenden Stamm. In Ungarn wird der Oberboden gesiebt und das verbleibende Rohsaatgut dann gereinigt.
In Bayern gibt es nur zwei Erntebestände im Raum Neustadt/Aisch und östlich von Bamberg. Sie wurden jedoch noch nie beerntet.
Bei einer Ernte muss auf jeden Fall von zehn Bäumen eines Bestands Saatgut eingesammelt werden, damit eine ausreichende genetische Vielfalt sichergestellt wird. Die Ernte erfolgt im Zeitraum November bis Februar durch Schütteln auf Netze bzw. durch Pflücken der Hülsen am stehenden Stamm. In Ungarn wird der Oberboden gesiebt und das verbleibende Rohsaatgut dann gereinigt.
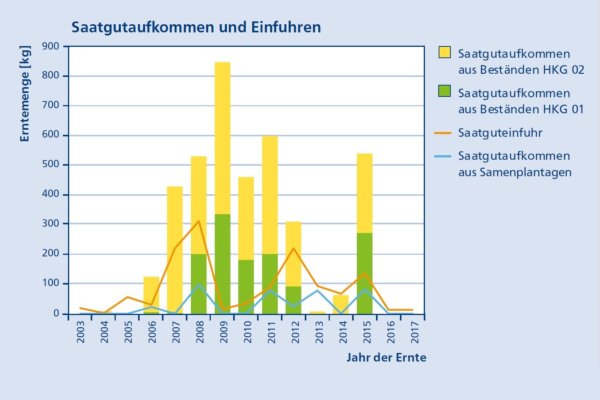 Zoombild vorhanden
Zoombild vorhanden
Abb. 6: Entwicklung des Saatgutaufkommens und der Einfuhren im Zeitraum 2003/17 (Grafik: LWF)
Zusätzlich gibt es deutschlandweit vier Samenplantagen mit insgesamt 2,2 ha. Die einzige süddeutsche Samenplantage liegt in Emmendingen bei Freiburg. Sie wurde 2006 auf einer Fläche von 0,3 ha angelegt und fruktifiziert bereits regelmäßig. Die jährlichen Ernteerträge in Mastjahren werden auf 20 – 70 kg Hülsen angeschätzt (Karopka 2020) (Abbildung 5).
Seit 2003 wurden in allen deutschen Zulassungsbeständen und Plantagen im Jahresmittel 260 kg Saatgut geerntet. Etwa 10 % des gesamten Saatgutaufkommens stammt aus Samenplantagen. Der Schwerpunkt der Ernte liegt in Brandenburg.
Zusätzlich wird regelmäßig Saat- und Pflanzgut vor allem aus Ungarn eingeführt. Der Saatgutimport beträgt etwa 20 % des Ernteaufkommens heimischer Bestände (Abbildung 6). Wegen der langen Lagerfähigkeit des Saatguts ist keine regelmäßige Ernte notwendig.
Seit 2003 wurden in allen deutschen Zulassungsbeständen und Plantagen im Jahresmittel 260 kg Saatgut geerntet. Etwa 10 % des gesamten Saatgutaufkommens stammt aus Samenplantagen. Der Schwerpunkt der Ernte liegt in Brandenburg.
Zusätzlich wird regelmäßig Saat- und Pflanzgut vor allem aus Ungarn eingeführt. Der Saatgutimport beträgt etwa 20 % des Ernteaufkommens heimischer Bestände (Abbildung 6). Wegen der langen Lagerfähigkeit des Saatguts ist keine regelmäßige Ernte notwendig.
Saatgutbehandlung und Anzucht
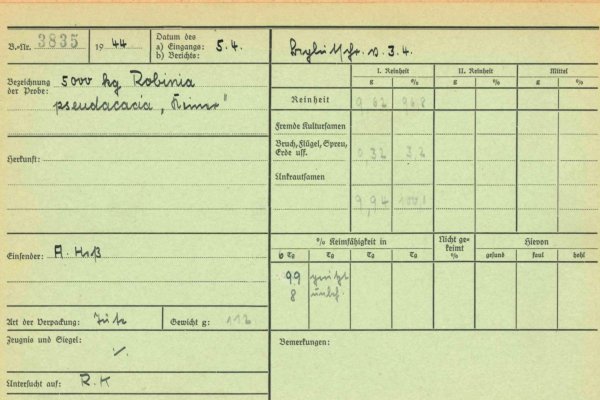 Zoombild vorhanden
Zoombild vorhanden
Abb. 7: Karteikarte mit Untersuchungsergebnis einer Robinien-Saatgutpartie zur Ermittlung der Reinheit und Keimfähigkeit aus dem Jahr 1944 (Quelle: Archiv, AWG)
Das Saatgut wird ab Mitte Oktober reif. In manchen Fällen bleibt es bis zum März am Baum hängen. In dieser Zeit baut sich durch die trockene Samenschale eine intensive Keimhemmung auf. In warmen Jahren mit einer hohen Wärmesumme ist diese Hartschaligkeit besonders ausgeprägt. Bei Lagertemperaturen unter 4 °C kann auf 5 % Wassergehalt heruntergetrocknetes Saatgut ohne Keimkraftverlust für zehn Jahre eingelagert werden.
Robiniensaatgut ist wegen des hohen Gehalts an dem Toxalbumin Robin giftig.
Robiniensaatgut überliegt in der Natur häufig. In Versuchen wurde bei im Boden vergrabenen Samen nach 20 Jahren noch eine Keimrate von 31 % beobachtet (Schubert o. A.) In Baumschulen wird die Keimhemmung des Saatguts wegen der sehr harten Samenschale durch Stratifikation abgebaut.
Verfahren sind die kurzzeitige Vorbehandlung in 40 °C heißem Wasser, das 24-stündige Einweichen in lauwarmem Wasser mit unmittelbar anschließender Aussaat oder das Ritzen der Samen mit Glasscherben in rotierenden Trommeln. Bewährt hat sich auch das Einweichen in Schwefelsäure für 10 bis 60 Minuten (Young 1992). Nach dieser Vorbehandlung keimen im Mittel etwa 70 – 80 % der Samen. Erste intensive Untersuchungen an Robiniensaatgut zur Ermittlung von Keimfähigkeit und Reinheit wurden in Bayern bereits Anfang der 1940er Jahre an unterschiedlichen Saatgutpartien durchgeführt (Abbildung 7).
Robiniensaatgut ist wegen des hohen Gehalts an dem Toxalbumin Robin giftig.
Robiniensaatgut überliegt in der Natur häufig. In Versuchen wurde bei im Boden vergrabenen Samen nach 20 Jahren noch eine Keimrate von 31 % beobachtet (Schubert o. A.) In Baumschulen wird die Keimhemmung des Saatguts wegen der sehr harten Samenschale durch Stratifikation abgebaut.
Verfahren sind die kurzzeitige Vorbehandlung in 40 °C heißem Wasser, das 24-stündige Einweichen in lauwarmem Wasser mit unmittelbar anschließender Aussaat oder das Ritzen der Samen mit Glasscherben in rotierenden Trommeln. Bewährt hat sich auch das Einweichen in Schwefelsäure für 10 bis 60 Minuten (Young 1992). Nach dieser Vorbehandlung keimen im Mittel etwa 70 – 80 % der Samen. Erste intensive Untersuchungen an Robiniensaatgut zur Ermittlung von Keimfähigkeit und Reinheit wurden in Bayern bereits Anfang der 1940er Jahre an unterschiedlichen Saatgutpartien durchgeführt (Abbildung 7).
Aus einem Kilogramm Hülsen lassen sich 150 – 330 g Saatgut gewinnen. In einem Kilo reinen Saatguts sind ca. 50.000 (30.000 – 100.000) Samen enthalten. Die Sämlingsausbeute liegt im Mittel jedoch nur bei 36.800 Sämlingen/kg reinem Saatgut (Krüssmann 1979). Im praktischen Baumschulbetrieb lassen sich aus einem Kilogramm gereinigtem Saatgut etwa 20.000 verkaufsfertige Sämlingspflanzen erzeugen.
Bei der Saatgutprüfung ergibt sich in Abhängigkeit von der Größe der Samen ein Tausendkorngewicht von durchschnittlich 20 g (7,3 bis 34 g). Südliche Herkünfte verfügen tendenziell über größere Samen (Erteld 1952).
Die Aussaat erfolgt Anfang Mai kurz vor dem Ende der Spätfrostperiode mit einer Saatgutmenge von 20 g/m2. Nach etwa vier Wochen laufen die Saaten auf, nach acht Wochen erreichen die Sämlinge bereits etwa 20 cm Höhe.
Da die Pflanzen im Herbst bereits ungefähr ein Meter hoch sind, werden im Regelfall im Frühjahr einjährige Sämlingspflanzen (Sortiment 30/50; 50/80; 80/120 cm) ausgebracht. Der Pflanzverband beträgt ca. 1,5 – 2 m x 1,5 m. Robinien sind besonders in der Jugend stark früh- und spätfrostgefährdet.
Die vegetative Vermehrung von Zuchtsorten erfolgt im Herbst durch die Gewinnung von 10 cm langen, mindestens 0,5 cm starken Wurzelstecklingen. Sie werden im Frühjahr in Beeten abgesteckt. Möglich ist auch die Steckholzvermehrung. Hierzu werden im Spätwinter etwa 20 cm lange, fingerstarke Abschnitte von Jahrestrieben gewonnen und vorzugsweise im Gewächshaus kultiviert.
Robinien zeigen nach Pflanzung häufig ein langsames Wachstum und unbefriedigende Stammformen. Es hat sich daher bewährt, diese Pflanzen nach einem Jahr auf den Stock zu setzen. Die Triebe sind danach sehr wüchsig und geradschaftiger.
Bei der Saatgutprüfung ergibt sich in Abhängigkeit von der Größe der Samen ein Tausendkorngewicht von durchschnittlich 20 g (7,3 bis 34 g). Südliche Herkünfte verfügen tendenziell über größere Samen (Erteld 1952).
Die Aussaat erfolgt Anfang Mai kurz vor dem Ende der Spätfrostperiode mit einer Saatgutmenge von 20 g/m2. Nach etwa vier Wochen laufen die Saaten auf, nach acht Wochen erreichen die Sämlinge bereits etwa 20 cm Höhe.
Da die Pflanzen im Herbst bereits ungefähr ein Meter hoch sind, werden im Regelfall im Frühjahr einjährige Sämlingspflanzen (Sortiment 30/50; 50/80; 80/120 cm) ausgebracht. Der Pflanzverband beträgt ca. 1,5 – 2 m x 1,5 m. Robinien sind besonders in der Jugend stark früh- und spätfrostgefährdet.
Die vegetative Vermehrung von Zuchtsorten erfolgt im Herbst durch die Gewinnung von 10 cm langen, mindestens 0,5 cm starken Wurzelstecklingen. Sie werden im Frühjahr in Beeten abgesteckt. Möglich ist auch die Steckholzvermehrung. Hierzu werden im Spätwinter etwa 20 cm lange, fingerstarke Abschnitte von Jahrestrieben gewonnen und vorzugsweise im Gewächshaus kultiviert.
Robinien zeigen nach Pflanzung häufig ein langsames Wachstum und unbefriedigende Stammformen. Es hat sich daher bewährt, diese Pflanzen nach einem Jahr auf den Stock zu setzen. Die Triebe sind danach sehr wüchsig und geradschaftiger.
Besonderheiten
Wegen des hohen Brennwerts von Robinienholz und des sehr raschen Jugendwachstums ist diese Baumart auch eine Alternative zum Anbau in sog. Energiewäldern auf trockneren, für Pappel nicht geeigneten Standorten. Voraussetzung ist jedoch eine Umtriebszeit von 15 – 20 Jahren, da nur durch den langen Umtrieb der spätere Kulminationspunkt des Zuwachses im Vergleich zu Pappelsorten ausgenutzt werden kann. Nachteilig ist das flächige Auflaufen von Wurzelbrut nach dem Rückschnitt. Wurzelbrut und hohe Härte des Holzes erschweren die nachfolgenden Ernten.
Da Robinien im Gegensatz zu zahlreichen anderen Baumarten auch auf trockenen Standorten eine Anbaualternative darstellen, können sie bei fortschreitender Klimaerwärmung zunehmend an Bedeutung gewinnen.
Da Robinien im Gegensatz zu zahlreichen anderen Baumarten auch auf trockenen Standorten eine Anbaualternative darstellen, können sie bei fortschreitender Klimaerwärmung zunehmend an Bedeutung gewinnen.
Herkunftsaspekte
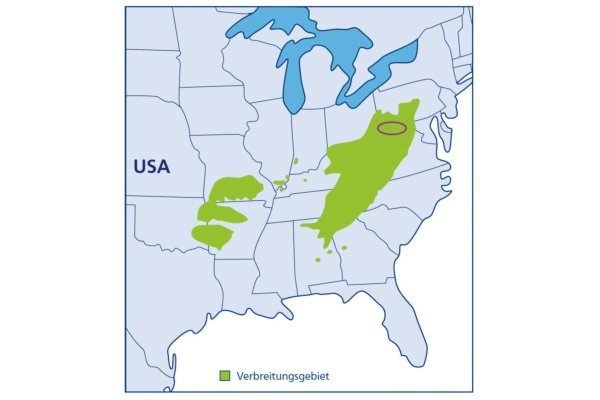 Zoombild vorhanden
Zoombild vorhanden
Abb. 8: Verbreitungsgebiet der Robinie in Nordamerika mit Vorkommensschwerpunkt der besten Herkünfte (violette Markierung) (Grafik: Quelle: https://fr.m.wikipedia.org/wiki/; 2020)
Robinie bildet in ihrer nordostamerikanischen Heimat aufgrund ihres Vorkommens in unterschiedlichen Höhenlagen und zwei isolierten Herkunftszonen unterschiedliche Klimarassen aus. Die besten Herkünfte stammen aus Höhenlagen zwischen 800 – 1.000 m NN in den Bundesstaaten Maryland und Virginia (Abbildung 8). Herkünfte aus südlichen und tiefliegenden Bereichen des natürlichen Verbreitungsgebiets fruktifizieren sehr häufig, bilden aber unbefriedigende Stammformen aus. Die Neigung zur Zwieselbildung ist herkunftsabhängig. Phototropisches Verhalten und Borkenstruktur sind ebenfalls genetisch fixiert.
Die als »Schiffsmastrobinie« bekannte Varietät Rectissima ist eine Zufallsselektion eines sehr wüchsigen, dickborkigen und besonders geradschaftigen Klons. Er stammt ursprünglich aus West Virginia und wurde um 1700 nach Long Island (Bundesstaat New York) verbracht und dort erfolgreich vegetativ weitervermehrt.
Wegen ihrer nördlichen Herkunft ist die »Schiffsmastrobinie« kältetoleranter und weniger spätfrostanfällig als südlichere Vorkommen. Diese Herkunft blüht kaum (Schenk 1939). Sie wird daher vegetativ über Wurzelstecklinge vermehrt. Phänotypisch ist sie an kürzeren, leicht nach oben gebogenen Dornen, grüngelben Blüten und deutlich eiförmigen Blättern erkennbar (Dengg 1994).
Ein Schwerpunkt der Züchtung besteht in Ungarn, wo zahlreiche aus vegetativ vermehrten Plusbäumen gezüchtete Sorten entstanden sind (Ewald et al. 1991, Ewald et al. 2001). Sehr geradschaftige Auslesebäume werden dort über Wurzelschnittlinge in Invitrokulturen angezogen. Sie werden als Cultivar Nyírségi und Kiskunsagi auf dem Markt angeboten und sind hinsichtlich Wuchsleistung, Form und Spätfrostresistenz deutlich besser geeignet für einen Wertholzanbau als generativ erzeugtes Pflanzmaterial (Schüler 2006).
Weitere für forstliche Zwecke geeignete ungarischen Sorten sind Jászkiséri und Penzesdombi sowie die in Amerika erfolgte Selektion Appalachia.
In einer griechischen Nachkommenschaftsprüfung wurde ein direkter Zusammenhang zwischen Individuen mit wenigen, aber großen Blättern und kleinen Dornen einerseits und Nachkommenschaften mit vielen kleinen Blättern und langen Dornen festgestellt (Dini-Papanastasi 2000).
Nachkommenschaftsprüfungen mit Robinie wurden in Deutschland erst im Zeitraum 1995/2004 angelegt, um die Vererbung des Merkmals Stammform zu untersuchen und um geeignetes Ausgangsmaterial für vegetative Selektionen zu lokalisieren (Liesebach 2004).
Die als »Schiffsmastrobinie« bekannte Varietät Rectissima ist eine Zufallsselektion eines sehr wüchsigen, dickborkigen und besonders geradschaftigen Klons. Er stammt ursprünglich aus West Virginia und wurde um 1700 nach Long Island (Bundesstaat New York) verbracht und dort erfolgreich vegetativ weitervermehrt.
Wegen ihrer nördlichen Herkunft ist die »Schiffsmastrobinie« kältetoleranter und weniger spätfrostanfällig als südlichere Vorkommen. Diese Herkunft blüht kaum (Schenk 1939). Sie wird daher vegetativ über Wurzelstecklinge vermehrt. Phänotypisch ist sie an kürzeren, leicht nach oben gebogenen Dornen, grüngelben Blüten und deutlich eiförmigen Blättern erkennbar (Dengg 1994).
Ein Schwerpunkt der Züchtung besteht in Ungarn, wo zahlreiche aus vegetativ vermehrten Plusbäumen gezüchtete Sorten entstanden sind (Ewald et al. 1991, Ewald et al. 2001). Sehr geradschaftige Auslesebäume werden dort über Wurzelschnittlinge in Invitrokulturen angezogen. Sie werden als Cultivar Nyírségi und Kiskunsagi auf dem Markt angeboten und sind hinsichtlich Wuchsleistung, Form und Spätfrostresistenz deutlich besser geeignet für einen Wertholzanbau als generativ erzeugtes Pflanzmaterial (Schüler 2006).
Weitere für forstliche Zwecke geeignete ungarischen Sorten sind Jászkiséri und Penzesdombi sowie die in Amerika erfolgte Selektion Appalachia.
In einer griechischen Nachkommenschaftsprüfung wurde ein direkter Zusammenhang zwischen Individuen mit wenigen, aber großen Blättern und kleinen Dornen einerseits und Nachkommenschaften mit vielen kleinen Blättern und langen Dornen festgestellt (Dini-Papanastasi 2000).
Nachkommenschaftsprüfungen mit Robinie wurden in Deutschland erst im Zeitraum 1995/2004 angelegt, um die Vererbung des Merkmals Stammform zu untersuchen und um geeignetes Ausgangsmaterial für vegetative Selektionen zu lokalisieren (Liesebach 2004).
Überblick zur Genetik
Wie der Großteil der Pflanzen und Bäume ist auch die Robinie diploid, d.h. sie besitzt einen doppelten Chromosomensatz (mit 2n = 22). Nur in Ausnahmefällen sind bei Robinie tetraploide Formen mit einem vierfachen Chromosomensatz, v.a. als Folge von Züchtung, gefunden worden. In den Chromosomen, die sich in jedem Zellkern befinden, ist die gesamte genetische Information gespeichert.
Genetische Analysen können dazu beitragen, Informationen über die Erbsubstanz zu gewinnen und damit Aufschluss über genetische Strukturen innerhalb von Populationen (z. B. genetische Variabilität) und über Differenzierungen zwischen Populationen zu geben. Des Weiteren können genetische Methoden, insbesondere DNA-Analysen, angewendet werden, um Individuen zu unterscheiden bzw. Klone zu identifizieren. Letzteres ist gerade bei der Robinie, die stark zu vegetativer (ungeschlechtlicher) Vermehrung neigt und auch für Zuchtformen (Sorten) vegetativ vermehrt wird, ein wichtigstes Instrument. So können natürliche klonale Strukturen innerhalb von Populationen, aber auch Robiniensorten (z. B. Gartenformen, Zuchtformen) eindeutig identifiziert und überprüft werden.
In einer Studie von Liesebach und Ewald 2012 wurden sechs Robinienbestände mittels sog. DNA-Mikrosatelliten an 13 DNA-Orten untersucht und ein genetischer Fingerabdruck für jede Robinie erstellt. Anhand dieser genetischen Daten ermittelten sie eine sehr geringe Wahrscheinlichkeit, dass Robinien zufällig den gleichen genetischen Fingerabdruck zeigen (1,67^–11 für nicht-verwandte Individuen, 1,37^–05 für Vollgeschwister). Mit dieser Identifizierungsmethode konnten sie in einigen Robinienbeständen starke vegetative Ausbreitung nachweisen.
Zur genetischen Variation innerhalb von Robinienbeständen in Deutschland bzw. Europa sind bisher vergleichsweise wenige Ergebnisse bekannt (z. B. Liesebach et al. 2004, Liesebach 2012, Liesebach und Ewald 2012). Genetische Untersuchungen mit Isoenzymen (Liesebach et al. 2004) zeigten beispielsweise eine geringe Variation zwischen verschiedenen Robinienbeständen in Ungarn, jedoch eine hohe Variation innerhalb der ungarischen Bestände. In deutschen Robinienbeständen verhielt es sich umgekehrt.
Die geringe Variation (genetische Diversität) innerhalb der deutschen Bestände ließ sich dabei durch die häufige Vermehrung über Wurzelbrut (klonale Strukturen) erklären, die durch waldbauliche Maßnahmen gefördert wurden. In brandenburgischen Beständen wurden z. B. Klone mit einer Flächenausdehnung von bis zu 0,5 ha beobachtet (Liesebach 2012). Die vergleichsweisen hohen genetischen Unterschiede zwischen deutschen Beständen (Liesebach et al. 2004) sind vermutlich auf die Begründung mit Pflanzgut aus unterschiedlichen Quellen zurückzuführen.
Die ungarischen Bestände entstanden meist aus Pflanzung und wurden waldbaulich in kurzen Umtrieben bewirtschaftet. Dadurch wurde die vegetative Vermehrung reduziert, was die höhere genetische Variation gegenüber den Ergebnissen in den deutschen Robinienbeständen erklärt. Da das Saatgut für die ungarischen Bestände meist aus einer bzw. aus nur wenigen Samenplantagen stammte, ist die genetische Differenzierung zwischen diesen eher gering.
Wegen der intensiven Vermehrung durch Wurzelbrut und die in der Forstpraxis häufige Verwendung von Sorten ist davon auszugehen, dass die Robinienvorkommen in Deutschland keine wie in den Ursprungsgebieten vergleichbar hohe genetische Vielfalt aufweisen. Um die Robinie beim Aufbau klimatoleranter Mischwälder angemessen zu beteiligen, muss jedoch sichergestellt werden, dass die Samenplantagen und regelmäßig beernteten Zulassungsbestände genetisch charakterisiert und bewertet werden, um eine ausreichend hohe genetische Diversität des Saatguts sicherzustellen. Bei der Begründung von Robiniensamenplantagen sollte vorab mit einer genetischen Prüfung ausgeschlossen werden, dass ausgewählte Plusbäume, die aus demselben Bestand stammen, zu einem Klon gehören (Naujoks et al. 2012).
Am Bayerischen Amt für Waldgenetik (AWG) wurden Methoden zur DNA-Analyse an Robinie 2015 etabliert und genetische Untersuchungen an einzelnen Individuen zur Sortenüberprüfung von Zuchtformen durchgeführt (z. B. für die Sorten »Sandraudiga«, »Monophylla «, »Semperflorens«). Diese Analysen bilden die Grundlage für die Erstellung eines Sortenkatasters basierend auf den genetischen Fingerabdrücken.
Genetische Analysen können dazu beitragen, Informationen über die Erbsubstanz zu gewinnen und damit Aufschluss über genetische Strukturen innerhalb von Populationen (z. B. genetische Variabilität) und über Differenzierungen zwischen Populationen zu geben. Des Weiteren können genetische Methoden, insbesondere DNA-Analysen, angewendet werden, um Individuen zu unterscheiden bzw. Klone zu identifizieren. Letzteres ist gerade bei der Robinie, die stark zu vegetativer (ungeschlechtlicher) Vermehrung neigt und auch für Zuchtformen (Sorten) vegetativ vermehrt wird, ein wichtigstes Instrument. So können natürliche klonale Strukturen innerhalb von Populationen, aber auch Robiniensorten (z. B. Gartenformen, Zuchtformen) eindeutig identifiziert und überprüft werden.
In einer Studie von Liesebach und Ewald 2012 wurden sechs Robinienbestände mittels sog. DNA-Mikrosatelliten an 13 DNA-Orten untersucht und ein genetischer Fingerabdruck für jede Robinie erstellt. Anhand dieser genetischen Daten ermittelten sie eine sehr geringe Wahrscheinlichkeit, dass Robinien zufällig den gleichen genetischen Fingerabdruck zeigen (1,67^–11 für nicht-verwandte Individuen, 1,37^–05 für Vollgeschwister). Mit dieser Identifizierungsmethode konnten sie in einigen Robinienbeständen starke vegetative Ausbreitung nachweisen.
Zur genetischen Variation innerhalb von Robinienbeständen in Deutschland bzw. Europa sind bisher vergleichsweise wenige Ergebnisse bekannt (z. B. Liesebach et al. 2004, Liesebach 2012, Liesebach und Ewald 2012). Genetische Untersuchungen mit Isoenzymen (Liesebach et al. 2004) zeigten beispielsweise eine geringe Variation zwischen verschiedenen Robinienbeständen in Ungarn, jedoch eine hohe Variation innerhalb der ungarischen Bestände. In deutschen Robinienbeständen verhielt es sich umgekehrt.
Die geringe Variation (genetische Diversität) innerhalb der deutschen Bestände ließ sich dabei durch die häufige Vermehrung über Wurzelbrut (klonale Strukturen) erklären, die durch waldbauliche Maßnahmen gefördert wurden. In brandenburgischen Beständen wurden z. B. Klone mit einer Flächenausdehnung von bis zu 0,5 ha beobachtet (Liesebach 2012). Die vergleichsweisen hohen genetischen Unterschiede zwischen deutschen Beständen (Liesebach et al. 2004) sind vermutlich auf die Begründung mit Pflanzgut aus unterschiedlichen Quellen zurückzuführen.
Die ungarischen Bestände entstanden meist aus Pflanzung und wurden waldbaulich in kurzen Umtrieben bewirtschaftet. Dadurch wurde die vegetative Vermehrung reduziert, was die höhere genetische Variation gegenüber den Ergebnissen in den deutschen Robinienbeständen erklärt. Da das Saatgut für die ungarischen Bestände meist aus einer bzw. aus nur wenigen Samenplantagen stammte, ist die genetische Differenzierung zwischen diesen eher gering.
Wegen der intensiven Vermehrung durch Wurzelbrut und die in der Forstpraxis häufige Verwendung von Sorten ist davon auszugehen, dass die Robinienvorkommen in Deutschland keine wie in den Ursprungsgebieten vergleichbar hohe genetische Vielfalt aufweisen. Um die Robinie beim Aufbau klimatoleranter Mischwälder angemessen zu beteiligen, muss jedoch sichergestellt werden, dass die Samenplantagen und regelmäßig beernteten Zulassungsbestände genetisch charakterisiert und bewertet werden, um eine ausreichend hohe genetische Diversität des Saatguts sicherzustellen. Bei der Begründung von Robiniensamenplantagen sollte vorab mit einer genetischen Prüfung ausgeschlossen werden, dass ausgewählte Plusbäume, die aus demselben Bestand stammen, zu einem Klon gehören (Naujoks et al. 2012).
Am Bayerischen Amt für Waldgenetik (AWG) wurden Methoden zur DNA-Analyse an Robinie 2015 etabliert und genetische Untersuchungen an einzelnen Individuen zur Sortenüberprüfung von Zuchtformen durchgeführt (z. B. für die Sorten »Sandraudiga«, »Monophylla «, »Semperflorens«). Diese Analysen bilden die Grundlage für die Erstellung eines Sortenkatasters basierend auf den genetischen Fingerabdrücken.
Zusammenfassung
Die Bedeutung von Robinie als wärmetolerante Baumart wird im Klimawandel zunehmen. Frühe Fruktifikation und ausgeprägte Neigung zur Wurzelbrut kennzeichnen sie als invasive Art. Da Robinien sehr starke Qualitätsunterschiede v. a. hinsichtlich der Schaftform aufweisen, darf Saat- und Pflanzgut nur aus amtlich zugelassenen Erntebeständen in Verkehr gebracht werden.
Das beste Saatgut kommt aus Samenplantagen bzw. von Herkünften der US-Staaten Virginia und Maryland. Aus dieser Region stammt auch die sog. Schiffsmastrobinie. Dieser Klon kann jedoch nur vegetativ vermehrt werden und ist im Handel nicht verfügbar.
Wegen der intensiven Vermehrung durch Wurzelbrut weisen deutsche Bestände eine geringe genetische Vielfalt auf. Eine genetische Untersuchung von Erntebeständen ist daher unverzichtbar, um anpassungsfähiges Vermehrungsgut bereitzustellen.
Das beste Saatgut kommt aus Samenplantagen bzw. von Herkünften der US-Staaten Virginia und Maryland. Aus dieser Region stammt auch die sog. Schiffsmastrobinie. Dieser Klon kann jedoch nur vegetativ vermehrt werden und ist im Handel nicht verfügbar.
Wegen der intensiven Vermehrung durch Wurzelbrut weisen deutsche Bestände eine geringe genetische Vielfalt auf. Eine genetische Untersuchung von Erntebeständen ist daher unverzichtbar, um anpassungsfähiges Vermehrungsgut bereitzustellen.
Literatur
- Biermayer, G. (1999): Fremdenfurcht unangebracht: Fremdländische Baumarten im Bayerischen Staatswald; LWFaktuell Nr. 20/1999
- Dengg, Ch. (1994): Biologie und Ökologie der Robinie (Robinia acacia L.) Diplomarbeit Forstwissenschaftliche Fakultät München. 165 S.
- Dini-Papanastasi (2000): Relation between growth and morphological trais and genetic parameters of Robinia pseudoacacia var. monophylla D.C. in northern Greece. Silvae Genetica 49, 1 (2000): 37–44
- Erteld, W. (1952): Wachstum und Ertrag der Robinie im Gebiet der DDR. In: Göhre, Die robinie und ihr Holz. Deutscher Bauernverlag Berlin
- Ewald, D. et al. (1992): Hat die Robinie in Brandenburg eine Zukunft? AFZ 47(12): 738–740
- Ewald, D. et al. (2001): Lassen sich selektierte Klone in waldbauliche Konzepte einbinden? AFZ 56(16): 816–818
- Hanover, J.W.; Merbathu, T.; Bloese P. (1991): Genetic improvement of black locust: a prime agroforesty species. Forestry Chronicle 67, 227–231
- Liesebach, H.; Yang, M.S.; Schneck V. (2004): Genetic diversity and differentiation in a black locust progeny test. Forest Genetics 11(2): 151–161. Arborea Publishers
- Liesebach, H. (2012): Genetische Charakterisierung von Robinienbeständen (Robinia pseudoacacia L.) in Deutschland mit nuklearen Mikrosatelliten-Markern: Erkenntnisse zu ihrer Bestandesbegründung. Beiträge aus der NW-FVA, Band 8
- Liesebach, H.; Ewald, E. (2012): Optimisation of a multiplex PCR assay of nuclear microsatellite markers for population genetics and clone identification in Robinia pseudoacacia L. Silvae Genetica 61, 4 5
- Mann, H.J.; Zimmermann, G. (1989): Unsere forstlichen Baumarten, Wirtschafts- und Forstverlag Euting KG.
- Naujoks G.; Ewald, D.; Ulrich, K.; Graeff, R. (2012): Bleibt die Züchtung der Robinie auf die vegetative Vermehrung beschränkt? AFZ-DerWald 16, 10–12
- Krüssmann, G. (1979): Die Baumschule 6. Auflage, Blackwell Wissenschaftsverlag Berlin – Wien
- Karopka, M. (2020): Mündliche Mitteilung vom 12.2. 2020, FVA Freiburg
- Schenk, C.A. (1939): Fremdländische Wald- und Parkbäume. Bd. 3 Die Laubhölzer Parey-Verlag Berlin
- Schubert J. (o. A.): Lagerung und Vorbehandlung von Saatgut wichtiger Baum- und Straucharten; Eberswalde-Finow; Landesanstalt für Ökologie, Bodenordnung und Forsten, NRW
- Schüler, S.; Weißenbacher, L.; Sieberer, K. (2006): Robinien für Energie- oder Wertholz – die Sorte macht’s! Forstzeitung 117(8): 8–9
- Schütt P. (2006): Robinia pseudoacacia. In: Schütt, Weisgerber, Schuck, Lang, Stimm, Roloff: Enzyklopädie der Laubbäume. Nikolverlag. Hamburg
- Young, J.A. (1992): Seeds of woody plants in North America; Disocorides Press, Portland