


RSS-Feed der Bay. Landesanstalt für Wald und Forstwirtschaft abonnieren
So verpassen Sie keine Neuigkeiten mehr. Unser RSS-Feed "Nachrichten der Bayerischen Landesanstalt für Wald und Forstwirtschaft" informiert Sie kostenlos über unsere aktuellen Beiträge.
Jörg Müller und Torben Hilmers
Katastrophen für den Menschen – Segen für die Biodiversität – LWF aktuell 127
Von der erhöhten Baumsterblichkeit durch zahlreiche Störungen gehen wichtige Impulse für Artenschutz und Biodiversität aus
Die steigende Mortalität von Waldbäumen stellt Manager und Gesellschaft vor große Herausforderungen. Allerdings leitet diese Mortalität für viele bedrohte Waldarten aktuell eine positive Trendwende ein. Daher sollten Chancen zum Schutz der Biodiversität jetzt nicht leichtfertig vergeben werden.
Frühe Sukzessionsphasen – das vergessene Habitat
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Störungen haben immer auch – was Biodiversität im Wald betrifft – ihr positiven Seiten. (Foto: M. Mößnang)
Verjüngung unter Schirm und gleichmäßige Durchforstungen sowie das Fehlen alter Wälder haben diese frühe Sukzessionsphase in Deutschland selten werden lassen (Hilmers et al. 2018). Auch im Naturschutz hat man diese artenreichste Phase stark vernachlässigt (Swanson et al. 2011).
Tote Bäume bieten wichtigen Lebensraum
 Zoombild vorhanden
Zoombild vorhanden
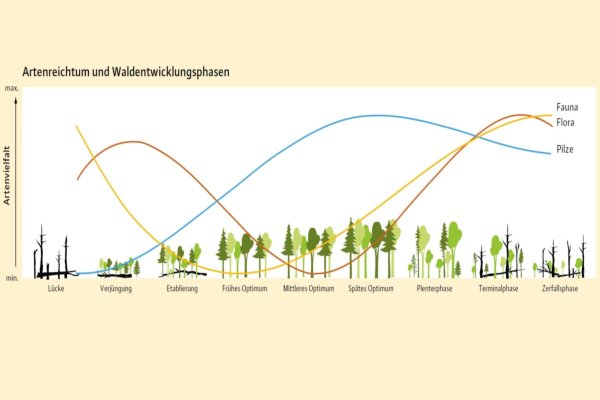
Abb. 2: Artenvielfalt im Wald entlang der Entwicklungsphasen (Grafik: LWF)
Die letzte Bundeswaldinventur hat sehr deutlich gezeigt, dass der Anstieg beim Totholz vor allem auf Nadelholz zurückzuführen ist: häufig Ergebnis unvollständiger Räumungen. So konnte in der Schweiz gezeigt werden, dass geräumte Windwurfflächen in Wirtschaftswäldern im Schnitt 75 m³/ha Totholz aufweisen (Priewasser et al. 2013).
Dieser Wert liegt weit über den üblich angestrebten Werten in genutzten Wäldern (Neft 2006; Müller & Bütler 2010). Darüber hinaus führen Störungen häufig – wenn auch nicht immer – zu sehr vielfältigen Totholzstrukturen, insbesondere dann, wenn betroffene Bäume zeitverzögert absterben, wie es häufig nach Feuer zu beobachten ist.
Hierzu wurde experimentell gezeigt, dass gerade die Totholzvielfalt der Schlüssel für eine hohe Insektenvielfalt ist (Seibold et al. 2016).
Störungen »reparieren« homogene Wirtschaftswälder
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Peltis grossa nach über 120 Jahren wider gesichtet. (Foto: L. Haselberger)
Ökologen aus den USA haben sogar die Theorie aufgestellt, dass Wälder nach großen Störungsereignissen komplex geboren werden können und auch nach 100 Jahren noch eine höhere Heterogenität im Gegensatz zum Wirtschaftswald aufweisen (Donato et al. 2012).
Störungen – Motor der Vielfalt
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: Zitronengelbe Tramete auf abgestorbenem Fichtenholz (Foto: C. Bässler)
Ein Vergleich befallener und intakter Fichtenbestände ergab über fast 20 taxonomische Artengruppen hinweg einen Zugewinn an Artenvielfalt durch den Borkenkäfer – und dies eben nicht nur bei Totholzbewohnern (Beudert et al. 2015). Dabei zeigte sich, dass im Bergwald viele Arten auf die frühen Sukzessionsstadien spezialisiert sind (Lehnert et al. 2013; Winter et al. 2017). Von der starken Auflichtung profitierte unter den Baumarten vor allem die Fichte und unter den Totholzkäfern vor allem die an Fichte gebundenen Arten (Müller et al. 2010).
Umgang mit Störungsflächen
- Räumung von Störungsflächen und anschließende Aufforstung zerstört die natürliche Heterogenität dieser so wichtigen und global seltenen Waldentwicklungsphase (Thorn et al. 2019). Gleichzeitig wird dadurch die nächste Generation homogener Waldbestände begründet (Donato et al. 2012).
- Arten, die deutschlandweit vor dem Aussterben stehen, können durch die aktuellen Störungen geschützt werden. Diese »Steilvorlage« sollte in Zeiten gesellschaftlicher Anstrengungen gegen den Rückgang von Insekten nicht ungenutzt bleiben.
- Besonnenes Abwarten, auch mehr als zehn Jahre nach Störungen, wäre in vielen Fällen naturschutzfachlich erstrebenswerter und deutlich ökonomischer. Rasche Wiederbewaldung wie vom Gesetz vorgesehen, steht hier im Wiederspruch zum Ziel, strukturreiche und gemischte Wälder im Klimawandel zu begründen.
- Inwieweit aktiv Baumarten erfolgversprechend eingebracht werden sollen, hängt von der Ausgangssituation und den Zielen des Eigentümers ab. Vielerorts, natürlich nicht immer, findet sich auf lichten Störungsflächen eine reiche Verjüngung heimischer Baum- und Straucharten ein. Mehrheitlich überlebt diese aktuell aber den selektiven Fraß hoher Rehpopulationen ohne Zaun nicht.
- Totholz auf Störungsflächen schafft naturnahe Lebensräume für viele Organismen. Wir sollten Waldbesitzer nicht dafür honorieren, wertvolle Lebensräume defizitär aus dem Wald zu schaffen, sondern wo immer möglich und auch ökonomisch sinnvoll, zu belassen.
- Forstschutzaspekte bei Totholz sind bisher nur bei der Fichte wissenschaftlich belegt. Dort ist genau zu prüfen, ob die Maßnahmen noch wirksam sind oder eher schon potenzielle Mopsfledermausquartiere vernichtet werden (Kortmann et al. 2018). Inzwischen gibt es auch verschiedene Alternativen, die kostengünstiger und schonender sind als zum Beispiel das Entrinden (Hagge et al. 2019).
Rückkehr der »Totgeglaubten« dank Störung
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Danosoma fasciata hat den Weg zurück in den Bayerischen Wald geschafft. (Foto: S. Thorn)
Um 1900 lebten in den naturnahen Wäldern des Bayerischen Waldes noch die seltenen Käferarten Lacon lepidopterus, Danosoma fasciata und Peltis grossa (Hennevogel 1905; Thiem 1906). Mit dem Einzug moderner Fichtenforsten verschwanden die Arten. Danosoma fasciata tauchte mit den Störungen nach 100 Jahren wieder auf. Heute trifft man ihn in diesen Flächen wieder regelmäßig an. Ein Exemplar besuchte gar einen Mitarbeiter des Nationalparks in seiner Badewanne am Waldrand.
Die Urwaldreliktart Peltis grossa kam erst 2019 zurück. Sie stammte wohl aus einer Prozessschutzfläche im Süden des Sumava Nationalparks, das sie als Refugium nutzte, ein kleines Naturschutzgebiet, ca. 40 km von der deutschen Grenze entfernt. Schlüsselhabitat sind wieder einmal starke Borkenkäferfichten mit Rotrandigem Baumschwamm. Lacon lepidopterus bleibt weiterhin verschollen. Eine gute Nachricht war seine Wiederentdeckung in Windwurfflächen im Nationalpark Sächsische Schweiz.
Aufräumen schadet der Artenvielfalt
Unberührte Störungsflächen werden von Experten heute als seltener eingeschätzt als alte Wälder. Eine Meta-Analyse hat gezeigt, dass Räumungen immer Gewinner und Verlierergruppen haben. Rund die Hälfte aller Waldartengruppen gehört hier aber zu den Verlierern, dies gilt insbesondere für Artengruppen mit Bindung an Totholz (Thorn et al. 2018). Eine neue Studie hat jetzt gezeigt, wie viel Störungsfläche belassen werden sollte, um einen bestimmten Anteil an Spezialisten der frühen Sukzessionsflächen erhalten zu können (Thorn et al. 2020).
Zusammenfassung
In Folge von Störungen entstehen unter anderem auch wichtige Habitate wie beispielsweise stehendes oder liegendes Totholz. Ungünstig für die Artenvielfalt wirken sich Räumungen der Störungsflächen aus. Am Schluss des Beitrages werden Empfehlungen genannt, wie mit Störungsflächen aus natuschutzfachlicher Sicht umgegangen werden soll.
- Bässler, C.; Müller, J. (2010): Importance of natural disturbance for recovery of the rare polypore Antrodiella citrinella Niemelä & Ryvarden. Fungal Biology 114: S. 129–133
- Beudert, B.; Bässler, C.; Thorn, S.; Noss, R.; Schröder, B.; Dieffenbach- Fries, H.; Foullois, N.; Müller, J. (2015): Bark beetles increase biodiversity while maintaining drinking water quality. Conservation Letters 8: S. 272–281
- Blaschke, M.; Helfer, W.; Ostrow, H.; Hahn, C.; Loy, H.; Bußler, H.; Krieglsteiner, L. (2009): Naturnähezeiger - Holz bewohnende Pilze als Indikatoren für Strukturqualität im Wald. Natur und Landschaft 84: S. 560–566
- Donato, D.C.; Campbell, J.L.; Franklin, J. F. (2012): Multiple successional pathways and precocity in forest development: can some forests be born complex? Journal of Vegetation Science 23, S. 576–584
- Grove, S. J. (2002): Saproxylic insect ecology and the sustainable management of forests. Annual Review of Ecology and Systematics 33: S. 1–23
- Hagge, J.; Leibl, F.; Müller, J.; Plechinger, M.; Soutinho, J.G.; Thorn, S. (2019): Reconciling pest control, nature conservation and recreation in coniferous forests. Conservation Letters 12:e12615
- Heidrich, L.; Levick, S.; Bae, S.; Magdon, P.; Serebryanyk, A.; Wöllauer, S.; Schall, P.; Ammer, C.; Nauss, T.; Krzystek, P.; Gossner, M.M.; Fischer, M.; Jung, K.; Kreft, H.; Heurich, M.; Doerfler, I.; Schulze, E.-D.; Hothorn, T.; Bässler, C.; Seibold, S.; Weisser, W.; Thorn, S.; Bauhus, J.; Müller, J. (2020): Revisiting MacArthur’s habitat-heterogeneity hypothesis on forest species diversity using airborne laser scanning. Nature Ecology & Evolution 36: 531
- Hennevogel, F. v. E. (1905): Zur Käferfauna des Böhmerwaldes. Verlag der Gesellschaft für Physiokratie in Böhmen, Prag
- Hilmers, T.; Friess, N.; Bässler, C.; Heurich, M.; Brandl, R.; Pretzsch, H.; Seidl, R.; Müller, J. (2018): Biodiversity along temperate forest succession. Journal of Applied Ecology 55: S. 2756–2766
- Kortmann, M.; Hurst, J.; Brinkmann, R.; Heurich, M.; Gonzalez, R.S.; Mueller, J.; Thorn, S. (2018): Beauty and the beast: how a bat utilizes forests shaped by outbreaks of an insect pest. Animal Conservation 21: S. 21–30
- Lehnert, L.W.; Bässler, C.; Brandl, R.; Burton, P.J.; Müller, J. (2013): Highest number of indicator species is found in the early successional stages after bark beetle attack. Journal for Nature Conservation 21: S. 97–104
- Lindenmayer, D.; Thorn, S.; Banks, S. (2017): Please do not disturb ecosystems further. Nature Ecology & Evolution 1: 0031
- Müller, J.; Bütler, R. (2010): A review of habitat thresholds for dead wood: a baseline for management recommendations. European Journal of Forest Research 129: S. 981–992
- Müller, J.; Noss, R.; Bussler, H.; Brandl, R. (2010): Learning from a »benign neglect strategy« in a national park: Response of saproxylic beetles to dead wood accumulation. Biological Conservation 143: S. 2559–2569
- Müller, J.; Noss, R.F.; Thorn, S.; Bässler, C.; Leverkus, A.B.; Lindenmayer, D. (2019): Increasing disturbance demands new policies to conserve intact forest. Conservation Letters 12:e12449
- Neft, R. (2006): Biotopbaum- und Totholzkonzept des Unternehmens Bayerische Staatsforsten (BaySF). LWF aktuell 55: S. 28–29
- Noss, R.F.; Lindenmayer, D.B. (2006): Special Section: The Ecological Effects of Salvage Logging after Natural Disturbance. Conservation Biology 20: S. 946–948
- Priewasser, K.; Brang, P.; Bachofen, H.; Bugmann H.; Wohlgemuth, T. (2013): Impacts of salvage-logging on the status of deadwood after windthrow in Swiss forests. European Journal of Forest Research 132: S. 231–240
- Seibold, S.; Bässler, C.; Brandl, R.; Büche, B.; Szallies, A.; Thorn, S.; Ulyshen, M.; Müller, J. (2016): Microclimate and habitat heterogeneity as the major drivers of beetle diversity in dead wood. Journal of Applied Ecology 53: S. 934–943
- Seibold, S.; Brandl, R.; Buse, J.; Hothorn, T.; Schmidl, J.; Thorn, S.; Müller, J. (2015): Association of the extinction risk of saproxylic beetles and the ecological degradation of forests in Europe. Conservation Biology 29: S. 382–390
- Seidl, R.; Schelhaas, M.-J.; Lexer, M. J. (2011): Unraveling the drivers of intensifying forest disturbance regimes in Europe. Global Change Biology 17: S. 2842–2852
- Senf, C.; Pflugmacher, D.; Yang, Z.Q.; Sebald, J.; Knorn, J.; Neumann, M.; Hostert, P.; Seidl, R. (2018): Canopy mortality has doubled in Europe›s temperate forests over the last three decades. Nature Communications 9
- Sommerfeld, A.; Senf, C.; Buma, B.; D›Amato, A.W.; Després, T.; Díaz-Hormazábal, I.; Fraver, S.; Frelich, L.E.; Gutiérrez, Á.G.; Hart, S.J.; Harvey, B.J.; He, H.S.; Hlásny, T.; Holz, A.; Kitzberger, T.; Kulakowski, D.; Lindenmayer, D.; Mori, A.S.; Müller, J.; Paritsis, J.; Perry, G.L.W.; Stephens, S.L.; Svoboda, M.; Turner, M.G.; Veblen, T.T.; Seidl, R. (2018): Patterns and drivers of recent disturbances across the temperate forest biome. Nature Communications 9: 4355
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; Dellasala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. (2011): The forgotten stage of forest succession: early-successional ecosystems on forest sites. Frontiers in Ecology and the Environment 9: S. 117–125
- Thiem, F.M. (1906): Biogeographische Betrachtung des Rachel. - zum Zwecke der Darlegung wie das Leben diesen Raum in vertikaler Richtung besetzt hat. Abhandlungen der Naturhistorischen Gesellschaft Nürnberg 16: S. 6–137
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.-Y.; Cobb, T.; Donato, D.C.; Durska, E.; Fontaine, J.B.; Gauthier, S.; Hebert, C.; Hothorn, T.; Hutto, R.L.; Lee, E.-J.; Leverkus, A.B.; Lindenmayer, D.B.; Obrist, M.K.; Rost, J.; Seibold, S.; Seidl, R.; Thom, D.; Waldron, K.; Wermelinger, B.; Winter, M.-B.; Zmihorski, M.; Müller, J. (2018): Impacts of salvage logging on biodiversity – a meta-analysis. Journal of Applied Ecology 55: S. 279–289
- Thorn, S.; Müller, J.; Leverkus, A.B. (2019): Forest dieback – Selling old problems as new solutions Science 365: 1388
- Thorn, S.; Chao, A.; Georgiev, K.B.; Müller, J.; Bässler, C.; Campbell, J.L.; Castro, J.; Chen, Y.-H.; Choi, C.-Y.; Cobb, T.P.; Donato, D.C.; Durska, E.; Macdonald, E.; Feldhaar, H.; Fontaine, J.B.; Fornwalt, P.J.; Hernández, R.M.; Hutto, R.L.; Koivula, M.; Lee, E.-J.; Lindenmayer, D.; Mikusiński, G.; Obrist, M.K.; Perlík, M.; Rost, J.; Waldron, K.; Wermelinger, B.; Weiß, I.; Żmihorski, M.; Leverkus, A.B. (2020): Estimating retention benchmarks for salvage logging to protect biodiversity. Nature Communications
- Turner, M.G.; Romme, W.H.; Tinker, D.B. (2003): Surprises and lessons from the 1988 Yellowstone fires. Frontiers in Ecology and the Environment 1: S. 351–358
- Winter, M.B.; Bässler, C.; Bernhardt-Römermann, M.; Krah, F.S.; Schaefer, H.; Seibold, S.; Müller, J. (2017): On the structural and species diversity effects of bark beetle disturbance in forests during initial and advanced early-seral stages at different scales. European Journal of Forest Research 136: S. 357–373
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren
- Jörg Müller
- Torben Hilmers