Karl-Heinz Mellert, Alwin Janßen und Muhidin Šeho
Anpassung an Klima und Boden bestimmt die Eignung von Herkünften - LWF aktuell 131
Auf der Suche nach geeigneten Saatguterntebeständen bringt das AWG mehrere wissenschaftliche Fachdisziplinen zusammen
In der Herkunftsforschung lag bislang der Fokus auf Mortalität, Vitalität, Wachstum und Qualität. Somit war der Forschungsansatz mehr auf bestimmte Phänotypen und weniger auf Standortfragen ausgerichtet. Wichtige genetische Prozesse finden jedoch entlang eines ökologischen Gradienten von feuchtkaltem zu warmtrockenem Klima statt. Daher wird in dem Forschungsprojekt »sensFORclim« der Einfluss der Genetik auf die Reaktion von Baumpopulationen entlang dieses ökologischen Gradienten untersucht.
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Studienmatrix der Buche: Grenzstandorte sind durch geringe Bodenwasserspeicherkapazität und marginales Klima gekennzeichnet.
In der Herkunftsforschung stehen neben der Mortalität und Vitalität traditionell Wachstum und Qualität von bewährten Herkünften im Zentrum der Aufmerksamkeit, da Herkunftsempfehlungen für die Forstpraxis neben dem Überleben auf die potenzielle Leistung und den erzielbaren Holzwert abzielen. Demensprechend waren die Studienansätze bisher auf bestimmte Phänotypen und weniger auf Standortfragen ausgerichtet. Entlang des ökologischen Gradienten von feucht-kaltem zu warm-trockenem Klima finden aber entscheidende demografische und genetische Prozesse statt (Fady et al. 2016). Traditionelle Herkunftsversuche sind aus den o.g. Gründen aber weniger geeignet, um die Klimatoleranz von Herkünften systematisch zu erforschen. Aktuell geforderte Alternativmaßnahmen wie »assisted gene flow« oder »assisted migration« bergen große Chancen, aber auch Risiken. Bevor derartige Transfers in großem Stil angegangen werden, ist primär zu prüfen, inwieweit lokale, in Deutschland und den einzelnen Bundesländern vorhandene Forstgenressourcen eine besondere Klimatoleranz aufweisen. Aus diesem Grund untersucht das interdisziplinäre Forschungsprojekt »sensFORclim« den Einfluss der Genetik auf die Reaktion von Baumpopulationen entlang ökologischer Gradienten (Abbildung 1), wobei der Klimagradient und die mögliche Anpassung eine zentrale Rolle spielen. Mögliche Effekte des Bodenwasserspeichers (Mellert et al. 2018b) und der Trophie (Mellert et al. 2018a) wurden im Studiendesign berücksichtigt. Institutionen aus vier Bundesländern (AWG, FVA Baden-Württemberg, Sachsenforst, Thüringenforst, Technische Universität München) arbeiten in »sens-FORclim« zusammen.
Studiendesign
In unserer Studie werden die drei Hauptbaumarten Fichte, Tanne und Buche untersucht. In diesem Beitrag zeigen wir auf, wie wir Herkünfte mithilfe von Nischenmodellen entlang ökologischer Gradienten reihen und in eine definierten Studienmatrix einordnen (Abbildung 1). Innerhalb des aufgespannten Umweltraumes werden angepasste bzw. anpassungsfähige Populationen mit modernen Methoden der Genetik, Baumphysiologie und Resilienzforschung identifiziert. Für die Genanalyse wurden 48 Bäume mit 30 m Abstand beprobt, woraus sich für die Untersuchungsbestände eine Mindestfläche von etwa 5 ha ergibt.
Klimatische Marginalität
 Zoombild vorhanden
Zoombild vorhanden
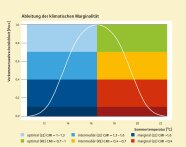
Abb. 2: Vorkommenswahrscheinlichkeit (Pocc) der Buche entlang des Temperaturgradienten (Sommer, Klimaperiode 1970–2000) mit Klassen der klimatischen Marginalität (CMI, farbige Flächen; vgl. Abbildung 3). Für die Darstellung in der Karte wurden marginale Extrembereiche (CMI < 0.1 bzw. > 1,9) durch dunklere Farbgebung hervorgehoben.
Die klimatische Vulnerabilität der Erntebestände wird in sensFORclim als klimatische Marginalität (CMI) (Mellert et al. 2015, 2016; Dorado et al. 2018) auf der Basis von Nischenmodellen mittels »Generalisierter Additiver Modelle« ermittelt. Die Baumvorkommen stammen aus dem Datensatz der LWF (2020) auf der Basis von ICP Forests-Daten (Mauri et al. 2017) und ergänzenden Quellen (Thurm et al. 2018). Um das klimatische Potenzial einer Baumart voll auszuschöpfen, wurde bereits bei einem Vorkommen innerhalb eines 16 x 16 km Rasters die Buche als präsent gewertet (Thurm et al. 2018). Als Klimadaten wurden WorldClim-Daten (WorldClim 2.1, Fick & Hijmans 2017) verwendet. Wie in Mellert et al. (2015) wurden als Basis für die Modellierung der Marginalität die Variablen BIO6 (Minimumtemperaturen im kältesten Monat), BIO10 (Mittlere Sommertemperaturen) und BIO18 (Sommerniederschlag) für die Klimaperiode 1970–2000 herangezogen.
Ursprünglich wurde die klimatische Marginalität von Standorten als »generelle Randigkeit« auf Ebene des lineraren Prädiktors (Logit) des logistischen Modells berechnet (Mellert et al. 2015). Hierbei wird nicht zwischen dem kalt-feuchten und dem warm-trockenen Nischenrand unterschieden. Bei der Klimaerwärmung ist jedoch vor allem der Abstand zum warm-trockenen Verbreitungsrand einer Baumart relevant, der auch als »Rear Edge« bezeichnet wird, weil sich die Bäume im Klimawandel daraus zurückziehen (Hampe & Petit 2005). Diesem Umstand wurde bei der weiteren Entwicklung Rechnung getragen (Mellert et al. 2016). In sensFORclim verwenden wir eine Variante der klimatischen Marginalität, bei der die Werte intuitiv auf der Ebene der Vorkommenswahrscheinlichkeit (Pocc) interpretiert werden können (Abbildung 2). Im Gegensatz zur Pocc, deren Intervall von 0 bis 1 reicht (Kurve in Abbildung 2), ist der Wertebereich von CMI 0–2 (siehe Hintergrundfarben in Abbildung 2). Dies erklärt sich folgendermaßen: Zur Ableitung dieses Maßes wird die Temperatur schrittweise soweit erhöht, bis Pocc am Verbreitungsrand praktisch auf 0 abfällt. Bei Vorkommen im Optimum gilt CMI = 1. Der Wert 2 wird für Vorkommen am Kältepol der Verbreitung (am Leading Edge) erreicht. Ein solches Vorkommen muss bei der klimatischen Annährung zum Rear Edge erst mal auf den »Optimumberg« (Weg auf der Wahrscheinlichkeitsskala ~1) ansteigen (Abbildung 2), um dort vom Gipfel wieder auf 0 abzusteigen. Die Summe der Wegstrecken im Wahrscheinlichkeitsraum bis zum Nischenrand ist demzufolge maximal 2. In sensFORclim interessieren jedoch vor allem Standorte, die dem Rear Edge zugewandt sind (rechte Seite der Grafik in Abbildung 2) und deren Marginalität dementsprechend unter 1 liegt. Klassen, aus denen Bestände ausgewählt wurden, liegen in den Bereichen optimal (CMI = 1,0–0,7; grün), intermediär (CMI = 0,7–0,4; gelb) und marginal (CMI = 0,4–0; rot). Bestände im Bereich des Leading Edge (CMI > 1) wurden vorerst nicht einbezogen.
Für die Untersuchungen in sensFORclim wurden die Bestände in Klassen der klimatischen Marginalität eingeteilt (optimale, intermediäre und marginale Klimaverhältnisse) und zwar unter einem moderaten Erwärmungsszenarium. Auf der Basis der aktuellen Marginalität (Klimaperiode 1970–2000) konnten im Untersuchungsgebiet nur für die Fichte klimatisch marginale Bestände (Marginalität < 0,4) ausgewiesen werden. Für die Tanne und die Buche, die ihr Verbreitungszentrum in Mitteleuropa haben, existieren für diese Klimaperiode keine marginalen Erntebestände. Deshalb wurde in sensFORclim ein Klimaerwärmungsszenarium ermittelt, bei dem die drei Marginalitätszonen bei allen drei Baumarten besetzt werden können. Wir testeten mehrere Temperaturerhöhungsszenarien und haben schließlich das Szenarium gewählt, das auf Daten der Klimaperiode 1970–2000 mit einer Temperaturerhöhung von + 2,5 °C basiert. Die entsprechende Marginalität bezeichnen wir mit CMI + 2,5. Folgende Aspekte waren für die Wahl dieses Szenariums ausschlaggebend:
- Es handelt sich dabei um ein mildes Szenarium, das sich nicht sehr stark vom Gegenwartsklima unterscheidet.
- Unter diesem Szenarium können im Untersuchungsgebiet Straten gebildet werden, welche die Zuweisung von künftig klimatisch marginalen Standorten zu den Erntebeständen bei allen Untersuchungsbaumarten erlauben (Abbildung 2 und 3).
- Ein räumliches Auseinanderdriften der klimatischen Zukunft von der Vergangenheit, wie es bei Anwendung der RCP-Szenarien auftreten kann, wird durch das einheitliche, für das gesamte Untersuchungsgebiet gültige Temperaturerhöhungsszenario vermieden.
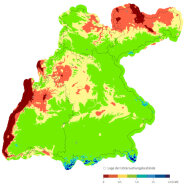
Die Konservierung der aktuellen räumlichen Klimamuster ist bei unserem Ansatz von Vorteil, da wir uns mit der genetischen Basis physiologischer Anpassungen von (autochthonen) Populationen befassen. Eine Entkoppelung der klimatischen Zukunft von der Vergangenheit wäre aber insbesondere für die retrospektiven Untersuchungen konzeptionell nachteilig. Die CMI + 2,5 der Standorte wird für alle vier Bundesländer flächendeckend berechnet (Abbildung 3).
Bodenwasserhaushalt
Neben dem Regionalklima im Untersuchungsgebiet wurde ein besonderes Augenmerk auf den lokalen Bodenwasserhaushalt gelegt. Zum einen um eine Überlagerung des Klimas durch Bodeneffekte durch Zuschusswasser zu vermeiden, zum anderen um den standörtlichen Grenzbereich zu definieren, der durch ungünstigen Bodenwasserhaushalt unter marginalen Klimaverhältnissen gekennzeichnet ist. Zur Vermeidung der genannten Überlagerungseffekte wurden grundsätzlich nur terrestrische Standorte ausgewählt. In den Klimastraten marginal bis optimal wurden durchschnittliche bis gute Standorte mit mittlerer bis hoher Wasserspeicherkapazität einbezogen. Die Wasserhaushaltstufe liegt hier i.d.R. im Bereich »mäßig frisch« bis »sehr frisch«. Lediglich im Optimalbereich, in dem definitionsgemäß weitgehend unlimitierte Wachstumsbedingungen herrschen, wurde Hangzugwasser toleriert. Grenzstandorte mit geringer Wasserspeicherkapazität bei marginalem Klima erreichen i.d.R. eine Wasserhaushaltsstufe von »mäßig trocken« oder ungünstiger. Diese Standorte stecken die standörtlichen Limits der jeweiligen Baumart zum Trockenen hin ab.
Bodentrophie
 Zoombild vorhanden
Zoombild vorhanden
Abb. 3: Klimatische Marginalität (CMI + 2,5) der Buche unter einem Szenarium von + 2,5 Grad Erwärmung gegenüber den gegenwärtigen Bedingungen (Klimaperiode 1970–2000).
Da unser Studiengebiet mehrere Bundesländer umfasst, haben wir für die Standortstrophie einen kompatiblen, länderübergreifenden Parameter verwendet, nämlich den Tiefengradienten der Basensättigung, kurz Basentyp (AK Standortskartierung 2016). Die Basensättigung ist eine zentrale bodenchemische Eigenschaft, da sie nicht nur die Verfügbarkeit der Makronährstoffe K, Ca und Mg beschreibt, sondern auch mit der Verfügbarkeit anderer wichtiger Nährstoffe zusammenhängt. Hohe Basensättigung in tief verwitterten Böden ist i.d.R. mit einer hohen N-Verfügbarkeit verbunden. Für die Baumernährung repräsentieren schwach-saure Böden das Optimum, da N, P sowie Fe, Mn und andere Mikronährelemente im Oberboden leicht verfügbar sind, während K, Ca und Mg als Makronährelemente im basenreichen Untergrund in großer Menge vorhanden sind (Kölling et al. 1996). Es lassen sich fünf Basentypen unterscheiden zwischen hoher Basensättigung des gesamten Profils (Typ 1) und durchgehend armer Basensättigung (Typ 5). Im Rahmen unseres Projektes haben wir hieraus drei Gruppen gebildet: basenreich (Basentypen 1 und 2), mittelbasisch (Basentyp 3) und basenarm (Basentypen 4 und 5). In Deutschland kommen Standorte vom Typ 1 und 2 sowie vom Typ 4 und 5 häufig in räumlich eng verzahnten Standortseinheiten bzw. Bodengesellschaften vor, weshalb sie für unsere Studie zusammengefasst wurden. Aufgrund der suboptimalen bis pessimalen Wuchsbedingungen im Marginal- und Grenzbereich konnten diese Straten nur zum Teil mit Erntebeständen besetzt werden. Um Populationen mit Genaustausch einzubeziehen, wurden Grenzbestände – soweit verfügbar – in der Nähe zu Marginalbeständen gesucht. Diese Anordnung der Untersuchungsbestände erleichtert es, Rückschlüsse auf etwaige Bodeneffekte zu ziehen.
Beobachtungen im Marginalbereich
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: Buchenbestand mit Trockenschäden aus dem Stratum »Grenzstandort, basenarm« (s. Abbildung 1). (Foto: Hoffmann, AWG)
Bei der aktuell laufenden Einrichtung der Untersuchungsbestände bestätigt sich die Stratifizierung des Klimaraumes und insbesondere die Definition des marginalen Bereichs insofern, dass es dort schwierig war, bei ungünstigerem Bodenwasserhaushalt Bestände ohne Trockenschäden zu finden. Dies gilt auch und gerade für die Buche, die bis vor Kurzem meist noch als »sichere Bank« im Klimawandel angesehen wurde. Im marginalen Bereich wiesen die Grenzbestände der Buche teilweise bereits deutliche Schäden auf (Abbildung 4), wohingegen die Bestände auf besseren Standorten augenscheinlich weniger betroffen waren. Verstärkte Trockenschäden im Marginalbereich werden auch von den dort anfallenden Schadholzmengen bestätigt. Die laufenden umfangreichen Untersuchungen in unserem systematischen Studiendesign dienen dazu, beobachtete Reaktionen von Baumpopulationen entlang der Standortgradienten zu quantifizieren und deren Bestimmungsfaktoren zu identifizieren. Besonders interessant ist die beobachtete Variabilität, die sich darin äußert, dass in der Nachbarschaft zu geschädigten Bäumen regelmäßig auch vitale Individuen bzw. Bestände vorkommen. Im Fokus stehen also jene Bestände, die sich vom beobachteten Trend einer abnehmenden Vitalität mit Annährung an den warm-trockenen »Rear Edge« positiv abheben. Denn solche Populationen können der Ausgangspunkt für die Nutzung heimischer Genressourcen in der Zukunft sein. Durch die Kombination moderner Methoden in einem systematischen Studiendesign in sensFORclim kann die Frage nach klimaplastischen Herkünften unserer heimischen Baumarten zielgerichtet und daher so effizient und zeitnah wie möglich beantwortet werden.
Das Projekt »sensFORclim«
Ziel des Projekts »sensFORclim« ist es, klimatolerante Saatguterntebestände der Baumarten Fichte, Buche und Tanne zu identifizieren und Vermehrungsgut für die Praxis verfügbar zu machen. Hierzu werden Saatguterntebestände in Sachsen, Thüringen, Baden-Württemberg und Bayern durch eine Kombination von ökologischen Nischenmodellen mit Methoden der Resillienzforschung sowie der Populationsgenetik untersucht. Das Projekt hat eine Laufzeit von 2020 bis 2023 und wird vom Bundesministerium für Ernährung und Landwirtschaft (BMEL) durch die Fachagentur für Nachwachsende Rohstoffe e.V. (FNR) gefördert.
Literatur
- AK Standortskartierung (2016): Forstliche Standortsaufnahme. IHW Verlag, Eching. 400 S. Bayerische Landesanstalt für Wald und Forstwirtschaft (LWF) (Hg.) (2020): Praxishilfe-Klima-Boden Baumartenwahl Band II, Freising, 124 S.
- Dorado‐Liñán, I., Piovesan, G., Martínez‐Sancho, E., Gea‐Izquierdo, G., Zang, C., Cañellas, I., ... & Fernández‐de‐Uña, L. (2019): Geographical adaptation prevails over species‐specific determinism in trees’ vulnerability to climate change at Mediterranean rearedge forests. Global change biology, 25(4), 1296-1314.
- Fady, B., Aravanopoulos, F. A., Alizoti, P., Mátyás, C., von Wühlisch, G., Westergren, M., ... & Zlatanov, T. (2016): Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. Forest Ecology and Management, 375, 66-75.
- Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: new 1‐km spatial resolution climate surfaces for global land areas. International journal of climatology, 37(12), 4302-4315.
- Hampe, A., & Petit, R. J. (2005): Conserving biodiversity under climate change: the rear edge matters. Ecology letters, 8(5), 461-467.
- Kölling, C., Hoffmann, M., & Gulder, H. J. (1996): Soil chemistry depth gradients as characteristic state variables of forest ecosystems. Zeitschrift für Pflanzenernährung und Bodenkunde, 159, 69-77.
- Mauri, A., Strona, G., & San-Miguel-Ayanz, J. (2017): EU-Forest, a high-resolution tree occurrence dataset for Europe. Scientific data, 4(1), 1-8.
- Mellert, K. H., Deffner, V., Küchenhoff, H., & Kölling, C. (2015): Modeling sensitivity to climate change and estimating the uncertainty of its impact: A probabilistic concept for risk assessment in forestry. Ecological Modelling, 316, 211-216.
- Mellert, K. H., Ewald, J., Hornstein, D., Dorado-Liñán, I., Jantsch, M., Taeger, S., ... & Kölling, C. (2016): Climatic marginality: a new metric for the susceptibility of tree species to warming exemplified by Fagus sylvatica (L.) and Ellenberg’s quotient. European journal of forest research, 135(1), 137-152.
- Mellert, K. H., Canullo, R., Mette, T., Ziche, D., & Göttlein, A. (2018a): Die klimatische Trockengrenze häufiger Baumarten hängt vom Bodennährstoffstatus ab. Schweizerische Zeitschrift für Forstwesen, 169(6), 323-331.
- Mellert, K. H., Lenoir, J., Winter, S., Kölling, C., Čarni, A., Dorado-Liñán, I., ... & Ewald, J. (2018b): Soil water storage appears to compensate for climatic aridity at the xeric margin of European tree species distribution. European Journal of Forest Research, 137(1), 79-92.
- Thurm, E. A., Hernandez, L., Baltensweiler, A., Ayan, S., Rasztovits, E., Bielak, K., ... & Falk, W. (2018): Alternative tree species under climate warming in managed European forests. Forest Ecology and Management, 430, 485-497.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren



 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden