LWF Wissen 86
Auswirkungen des Waldumbaus mit Buche auf strukturelle Vielfalt und Biodiversität
von Thomas Kudernatsch und Bastian Schaue
Zusammenfassung: Im Rahmen des Projekts L59 wurden die Effekte des Waldumbaus von Fichtenreinbeständen zu Fichten-Buchen-Mischbeständen auf Waldstruktur und Biodiversität anhand drei verschiedener Bewirtschaftungssysteme (Fichtenreinbestände, Fichten-Buchen-Mischbestände mit geringem bzw. hohem Buchenanteil) analysiert. Die Untersuchungen wurden im Ebersberger Forst, einem standörtlich homogenen Waldkomplex im Süden Deutschlands, durchgeführt. Dabei zeigte sich, dass der Umbau von Fichtenreinbeständen in Fichten-Buchen-Mischbestände dazu beiträgt, die strukturelle Vielfalt, Diversität und damit Funktionalität der Wälder zu erhöhen. Die Beimischung der Buche wirkt sich v. a. auf die Zusammensetzung der Artengemeinschaften (Beta-Diversität) aus, weniger auf deren Artenreichtum (Alpha- bzw. Gesamtdiversität). Die bestmögliche Förderung der Gesamtvielfalt auf Landschaftsebene kann durch die Kombination aller drei Bewirtschaftungssysteme erreicht werden. Im alpennahen Ebersberger Forst mit seinen vergleichsweise hohen Niederschlägen kann daher auch der Erhalt einzelner Fichtenreinbestände eine ökologisch sinnvolle Option sein.
Der Umbau der immer noch großflächig vorhandenen, nicht standortgemäßen reinen Nadelwälder in naturnähere, standortgerechte und stabile Mischwälder ist eine der wichtigsten Aufgaben einer modernen, zukunftsorientierten Forstwirtschaft in Deutschland (von Teuffel et al. 2005). Ein wesentliches Ziel des Waldumbaus ist es, die Stabilität und Elastizität der Waldökosysteme gegenüber biotischen und abiotischen Störungen zu erhöhen. Weiterhin wird durch den ökologischen Waldumbau aber auch eine Erhöhung der Biodiversität in den Waldökosystemen erwartet, wobei hierunter nicht nur die Vielfalt der Tier, Pilz und Pflanzenwelt, sondern auch die strukturelle Vielgestaltigkeit verstanden wird. Tatsächlich gibt es aber nur wenige Studien, die die Auswirkungen von Waldumbaumaßnahmen auf Biodiversität und strukturelle Vielfalt (insbesondere auf Ebene der Landschaft) tatsächlich untersucht haben (von Teuffel et al. 2005; Heinrichs et al. 2019).
Um die diesbezügliche Wissenslücke zu verkleinern, wurde das aus Mitteln der Bayerischen Forstverwaltung geförderte Projekt L59 durchgeführt. In diesem Vorhaben wurden die Effekte des Waldumbaus auf Waldstruktur und Biodiversität anhand verschiedener Bewirtschaftungssysteme (Fichtenreinbestandswirtschaft vs. Umbau von Fichtenreinbeständen in Fichten-BuchenMischbestände mit geringem bzw. hohem Laubholzanteil) analysiert. Um die Wirkungen der Waldbewirtschaftung auch auf Ebene der Landschaft erfassen zu können, wurden pro Bewirtschaftungssystem jeweils mehrere Flächen untersucht, die alle relevanten Bestandesphasen repräsentieren (von der initialen Verjüngung über die Stangenholz- bis zur Baumholzphase). Auf den Probeflächen, die sich allesamt im standörtlich sehr homogenen Ebersberger Forst befinden, fanden nach standardisierten Methoden Kartierungen der Waldstruktur, des Humus sowie verschiedener, bioindikatorisch aussagekräftiger Artengruppen statt. Durch diese wissenschaftliche Fallstudie kann somit das begrenzte Wissen um die Auswirkungen von Waldumbau auf die verschiedenen Ebenen der Biodiversität mittels eines innovativen Versuchsansatzes erweitert werden.
Fragestellung
Im Rahmen des Projekts wurden insbesondere folgende Hypothesen überprüft:
- Der Umbau von Fichtenreinbeständen führt zu einer größeren Struktur und Nischenvielfalt der Wälder.
- Der Umbau von Fichtenreinbeständen führt zu einer Erhöhung der Alpha-, Beta- und Gesamt-Diversität.
- Der Effekt der Umbaumaßnahmen auf Struktur- und Artenreichtum/-zusammensetzung steigt mit zunehmenden Laubholzanteil der Bestände an
- Eine Kombination verschiedener Bewirtschaftungssysteme innerhalb einer Landschaft beeinflusst die Biodiversität positiv.
Untersuchungsgebiet
Der Ebersberger Forst liegt etwa 25 Kilometer östlich von München in den Wuchsgebieten 13 »Schwäbisch-Bayerische Schotterplatten und Altmoränenlandschaft« und 14 »Schwäbisch-Bayerische Jungmoräne und Molassevorberge«. Der überwiegende Teil (und auch das Untersuchungsgebiet im engeren Sinne) ist dem Teilwuchsbezirk 13.2/1 Südliche Münchner Schotterebene zuzuordnen. Dieser weist durch die Nähe zum Alpenrand ein niederschlagsreiches Klima auf. Nach PIK (2009) wird für das FFHGebiet »Ebersberger und Großhaager Forst« für den Referenzzeitraum 1961 – 1990 eine Jahresmitteltemperatur von 7,3 °C bei 971 mm Jahresniederschlag angegeben.
Durch die hohen Niederschläge sind große Anteile der Standorte tiefergehend entbast, insbesondere im Bereich älterer Schotter- und der Decklehme (Jerz 1993). Natürliche Leitgesellschaft ist daher der bodensaure Hainsimsen-Buchenwald (Luzulo-Fagetum), mit Buche als Haupt- und Tanne als Nebenbaumart. Die Fichte ist in diesem Wuchsraum als eingebürgerte Baumart anzusehen (Walentowski et al. 2001).
Während der Ebersberger Forst von Natur aus also ein buchendominierter Wald wäre, ist das heutige Erscheinungsbild noch stark durch die seit dem 19. Jahrhundert auf großer Fläche verbreiteten Fichtenforste geprägt. Diese wurden begründet, um die seinerzeit devastierten Wälder wieder in Bestockung zu bringen und den durch die einsetzende Industrialisierung zunehmenden Holzbedarf zu decken. In den 1890er Jahren wurden durch Nonnenfalter-Kalamitäten und zyklonartige Wirbelstürme ausgedehnte Kalamitätsflächen verursacht. Die großen Kahlflächen konnten damals aufgrund von Spätfrostereignissen, starker Vergrasung und damit einhergehendem Mäuseschäden sowie Wildverbiss wiederum nur mit Fichten erfolgreich bestockt werden, die Laubbäume fielen weitgehend aus (Sponholz 1975). Seit Mitte/Ende des letzten Jahrhunderts wird versucht, durch gezielten Waldumbau im Schutz der Nadelholz-Altbestände wieder einen naturnäheren Laub-/Mischwald aufzubauen, um zukünftigen Sturmschäden und Insektenkalamitäten vorzubeugen. Die im Rahmen des Voranbaus bedeutsamste Baumart innerhalb des Ebersberger Forstes ist dabei die Buche.
Versuchsdesign
Im Rahmen des Projekts wurden die Effekte des Waldumbaus auf Biodiversität und Waldstruktur anhand eines Vergleichs der folgenden drei Bewirtschaftungssysteme analysiert:
- Fichtenreinbestandswirtschaft (»Z« – Zero: Buchenanteil = 0 %)
- Umbau von Fichtenreinbeständen in Fichten-Buchen-Mischbestände mit geringem Buchen-Anteil (»L« – Low: Buchenanteil = 5 – 10 %)
- Umbau von Fichtenreinbeständen in Fichten-Buchen-Mischbestände mit hohem Buchen-Anteil (»H« – High: Buchenanteil = 30 – 50 %)
Da der Waldumbau im Ebersberger Forst seit umgerechnet einem halben Jahrhundert aktiv betrieben wird, war es ferner möglich, verschiedene Stadien des Umbaus bzw. der Waldentwicklung gleichzeitig zu betrachten, wodurch insgesamt eine zuverlässigere Aussage zur Wirkung der Waldbewirtschaftung auf Ebene der Landschaft möglich ist (Ammer et al. 2017). Um diesem Aspekt Rechnung zu tragen, wurden daher innerhalb eines jeden Bewirtschaftungssystems folgende Entwicklungsstadien (Nutzungsarten) berücksichtigt:
- Verjüngungsnutzung (»VJ«): initiale Verjüngung
- Jungdurchforstung (»JD«): Stangenholzstadium
- Altdurchforstung (»AD«): Baumholzstadium
Die daraus resultierenden neun Varianten wurden durch jeweils sechs ein Hektar große Probeflächen repräsentiert (Abb. 1). Die Auswahl der insgesamt 54 Flächen basierte auf aktuellen Inventur- bzw. Forsteinrichtungsdaten der Bayerischen Staatsforsten. Selektiert wurde nach Nutzungsart, Buchenanteil im Altbestand bzw. der Vorausverjüngung und Bestandesgröße. Zusätzlich wurde ein Anteil von maximal 5 % »Fremdbaumarten« (alle Baumarten außer Fichte und Buche) toleriert.
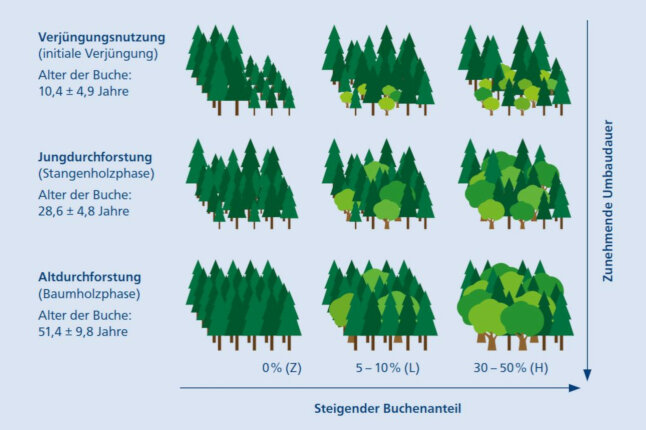
Abb. 1: Versuchsdesign: Je Bewirtschaftungssystem (links: Fichtenreinbestandswirtschaft, Mitte: Umbaubetrieb mit geringem Buchenanteil, rechts: Umbaubetrieb mit hohem Buchenanteil) und Nutzungsart wurden jeweils sechs Flächen untersucht, woraus ein Gesamtumfang von insgesamt 54 Probeflächen resultierte. (© LWF)
Charakterisierung der Struktur- und Artenvielfalt
Um zu untersuchen, wie sich Waldumbau auf die strukturelle Vielfalt auswirkt, wurde für jede Probefläche ein Heterogenitätsindex (Storch et al. 2018) errechnet. Die einzelnen Parameter, die in den Index eingeflossen sind, stammen aus der durchgeführten Bestandesinventur (lebender Bestand, Totholz, Verjüngung), Mikrohabitatkartierung sowie Humusansprache (vgl. Tabelle 1). Für jede Probefläche kann somit ein Heterogenitätswert ermittelt werden, der zwischen 0 und 1 liegt (0 = geringstmögliche, 1 = höchstmögliche strukturelle Vielfalt).
Die Ermittlung der Artenvielfalt und -zusammensetzung auf den Flächen erfolgte mittels auf die jeweilige Artengruppe abgestimmter Methoden. Um die Waldbodenpflanzen zu erfassen, wurden jeweils zwei pflanzensoziologische Aufnahmen je Probefläche durchgeführt. Zur Kartierung der Pilzfruchtkörper wurde jede Fläche dreimal begangen, um der Saisonalität der verschiedenen Pilzarten gerecht zu werden. Die Vogelarten wurden mittels kombinierter Punkt-Stopp-Revierkartierung dokumentiert. Die Kartierung erfolgte in drei Durchgängen zwischen Ende März und Anfang Juni. Bei den Arthropoden kamen Bodenfallen bzw. Kreuzfensterfallen zur Anwendung. Die Fangperiode dauerte von Anfang Mai bis Ende September. Die Artbestimmung erfolgte sowohl klassisch als auch durch Metabarcoding (Fänge aus den Flugfensterfallen).
Im Rahmen der Auswertung wurden verschiedene Ebenen der Diversität unterschieden. So wurde für jedes Bewirtschaftungssystem sowohl die Alpha-, Beta- als auch die Gesamtdiversität ermittelt. Unter Alpha-Diversität wird im Folgenden die lokale Artenvielfalt, also die Diversität der einzelnen Probeflächen verstanden (ausgedrückt über die Anzahl der auf den einzelnen Probeflächen vorkommenden Arten). Die Beta-Diversität hingegen gibt Auskunft darüber, wie sehr sich die einzelnen Probeflächen eines Kollektivs (hier Bewirtschaftungssystem) hinsichtlich ihrer Artenzusammensetzung unterscheiden. Je höher die Beta-Diversität, desto höher ist die Unähnlichkeit der Artenzusammensetzung bzw. der Artenwechsel zwischen den einzelnen Aufnahmeflächen. Die Gesamt-Diversität schließlich gibt an, wie viele Arten – über alle Probeflächen eines Kollektivs hinweg – insgesamt erfasst wurden.
Die auf den einzelnen Probeflächen dokumentierten Artnachweise wurden ferner dazu genutzt, die potenzielle (also auf Landschaftsebene maximal mögliche) Gesamtartenvielfalt der einzelnen Bewirtschaftungssysteme bzw. verschiedener Bewirtschaftungssystem-Kombinationen (ZL, ZH, LH, ZLH) abzuschätzen. Dazu wurden sog. Species Accumulation Curves berechnet.
Strukturvielfalt nimmt waldumbaubedingt zu
 Zoombild vorhanden
Zoombild vorhanden
Abb. 2: Boxplots der für die einzelnen Probeflächen errechneten Heterogenitätsindices, unterteilt nach Bewirtschaftungssystem.
Die Flächen mit hohem Buchenanteil (H) zeigten mit durchschnittlich 0,41 einen signifikant höheren Heterogenitätswert als die Z und L-Flächen (mittlerer Heterogenitätswert 0,29 bzw. 0,32). Zwischen Z und L-Flächen konnte kein signifikanter Unterschied nachgewiesen werden (Abb. 2).
Tatsächlich konnte im Rahmen unserer Studie also gezeigt werden, dass durch die Einbringung von Buche in Fichtenreinbestände eine Erhöhung der strukturellen Vielfalt zu erreichen ist. Im vorliegenden Fall beruht der Anstieg v. a. auf einer Zunahme der Baumartenvielfalt sowie einer stärker gestuften, arten- und individuenreicheren Verjüngungsschicht. Positiv auf die Heterogenität wirken sich ferner eine höhere Variabilität der Humusauflage sowie ein größeres Angebot an Mikrohabitaten aus (Tab. 1).
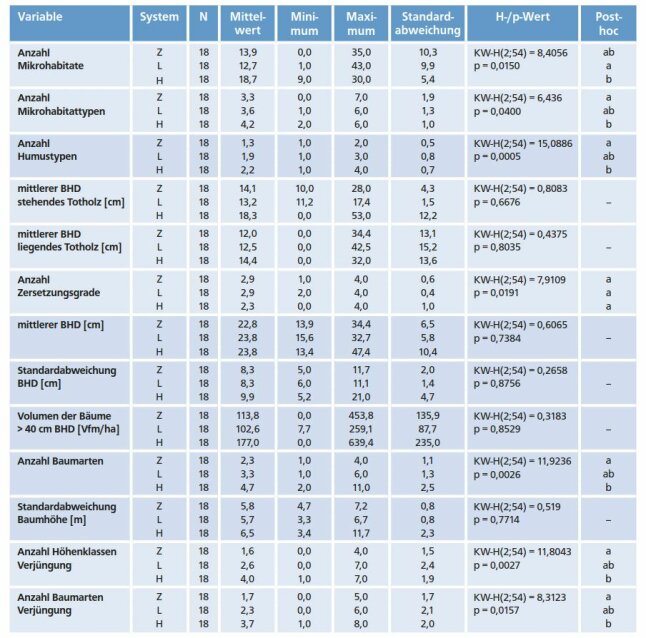
Tab. 1: Ausprägung der in den Heterogenitätsindex eingegangenen Strukturparameter in den drei Bewirtschaftungssystemen.
Auswirkungen auf die Artenvielfalt
Über alle untersuchten Artengruppen hinweg ist die Alphadiversität auf den Flächen mit geringem Buchenanteil (L) mit durchschnittlich 249 Arten am höchsten, gefolgt von den H-Flächen mit 233 und den Z-Flächen mit 225 Arten. Die Artenzahlen der Fichtenreinbestände und der Flächen mit geringem Buchenanteil unterscheiden sich dabei signifikant, während sich die H-Flächen hinsichtlich der Alphadiversität nicht von den anderen beiden Bewirtschaftungssystemen absetzen. Eine pauschale Erhöhung der lokalen Artenvielfalt mit steigenden Buchenanteilen war also nicht zu beobachten (Abb. 3).
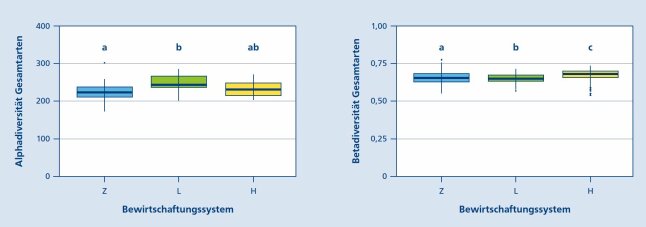
Abb. 3: links: Boxplots der auf den einzelnen Untersuchungsflächen nachgewiesenen Gesamtartenzahlen, unterteilt nach Bewirtschaftungssystem; rechts: Boxplots der Betadiversität (Jaccard-Index) innerhalb der verschiedenen Bewirtschaftungssysteme. Unterschiedliche Buchstaben markieren signifikante Unterschiede zwischen den Bewirtschaftungssystemen. (© LWF)
Unterschiede zwischen den Bewirtschaftungssystemen gibt es auch hinsichtlich der Beta-Diversität, die mittels des JaccardIndex quantifiziert wurde. Diese war in den H-Flächen signifikant höher als in den anderen beiden Systemen. Dies bedeutet, dass der Artenwechsel zwischen den einzelnen Probeflächen innerhalb der H-Variante am größten ist (Abb. 3).
 Zoombild vorhanden
Zoombild vorhanden
Abb. 4: Venn Diagramm der Gesamtdiversität der drei Bewirtschaftungssysteme. (© LWF)
Die Gesamtdiversität schließlich stieg waldumbaubedingt leicht an. Während in den Fichtenreinbeständen (Z) insgesamt 949 Arten nachgewiesen wurden, waren es auf den Umbauflächen 974 (L) bzw. 1.003 Arten (H). Jedes Bewirtschaftungssystem war dabei durch »exklusive«, also ausschließlich dort nachgewiesene Arten gekennzeichnet. Der höchste Anteil exklusiver Arten wurde in den Flächen mit hohem Buchenanteil gefunden (17 %), gefolgt von Fichtenreinbeständen (14 %) und Beständen mit geringem Buchenanteil (13 %; Abb. 4).
Dass sich die einzelnen Artengruppen hinsichtlich ihrer Reaktion zum Teil deutlich unterscheiden, geht aus Tabelle 2 hervor. Diese gibt einen vereinfachten Überblick über die Alpha und Betadiversität der untersuchten Artengruppen in den drei Bewirtschaftungssystemen.
Auffällig ist, dass die Beimischung der Buche in den wenigsten Fällen eine Erhöhung der lokalen Artenvielfalt nach sich zieht. So können für sechs der acht Artengruppen keine waldumbaubedingten Unterschiede der Alphadiversität aufgezeigt werden. Lediglich bei den Pilzen ist der Artenreichtum in den Fichten-Buchen-Mischbeständen nachweislich höher als in den Fichtenreinbeständen. Die lokale Diversität der Käfer zeigte eher einen negativen Trend.
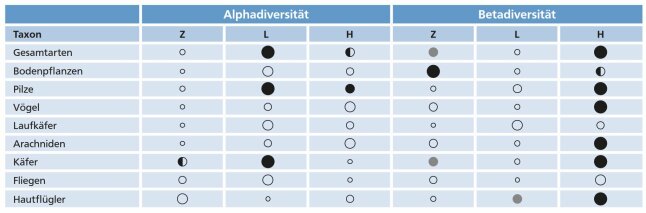
Tab. 2: Alpha- und Betadiversität der untersuchten Artengruppen in den drei Bewirtschaftungssystemen. Je größer die Kreise, desto höher sind die Mittelwerte der gefunden Alpha- und Betadiversitätswerte. Verschiedene Kreisfarben zeigen signifikante Unterschiede zwischen den verschiedenen Bewirtschaftungssystemen an. (© LWF)
Deutlich anders ist es bei der Betadiversität. Hier weisen die Flächen mit hohem Buchenanteil vielfach die höchsten Diversitätswerte auf. Dies gilt für fünf der acht untersuchten Artengruppen. Alles in allem führte der Waldumbau mit Buche also weniger zu einem Anstieg der Artenzahlen auf den Flächen als vielmehr zu einem erhöhten Artenwechsel zwischen den einzelnen Probeflächen innerhalb eines Bewirtschaftungssystems. Zu einer Zunahme der Betadiversität kommt es aber meist erst, wenn die Buchenbeimischung einen Anteil von 20 bis 30 % übersteigt.
Kombination der Bewirtschaftungssysteme
 Zoombild vorhanden
Zoombild vorhanden
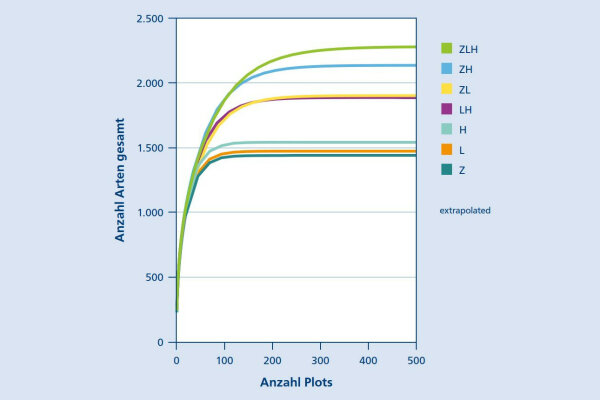
Abb. 5: Extrapolation der Gesamtartenzahlen mittels Species Accumulation Curves. Betrachtet werden die jeweiligen Einzelsysteme sowie deren Kombinationen. (© LWF)
Durch Extrapolation mittels Species Accumulation Curves ist es möglich, die potenzielle Gesamt-Diversität in einem Untersuchungsgebiet zu ermitteln. Dieses Verfahren wurde hier genutzt, um abzuschätzen, welche Gesamtdiversität innerhalb der einzelnen Bewirtschaftungssysteme (Z, L, H), bei Kombination von zwei Systemen (ZL, ZH, LH) bzw. bei gleichzeitigem Vorhandensein aller drei Systeme (ZLH) zu erwarten wäre.
Dabei ist eine deutliche Abstufung zu erkennen: So weisen die Einzelsysteme die niedrigste zu erwartende Artenvielfalt auf, wobei die Unterschiede zwischen den drei Systemen vergleichsweise gering sind. Eine Kombination von zwei Systemen führt hingegen schon zu wesentlich höheren Gesamtdiversitäten auf Landschaftsebene, insbesondere, wenn Z- und H-System miteinander kombiniert werden.
Die mit Abstand höchste Diversität kann jedoch bei Kombination aller drei Bewirtschaftungssysteme erreicht werden (Abb. 5).
Diskussion
Durch die Veränderung der Struktur eines homogenen Habitats, wie zum Beispiel bei der Einbringung einer neuen Baumart in einen Reinbestand, ist auch immer mit einer Veränderung der verfügbaren Nischen zu rechnen. In der Regel wird davon ausgegangen, dass gemischte Bestände eine größere Bandbreite und ein höheres Angebot an Nischen und damit Habitaten aufweisen (Böhme 2001). Tatsächlich konnte im Rahmen unserer Studie gezeigt werden, dass mit zunehmenden Buchenanteil auch die Strukturvielfalt anstieg, wobei sich die H-Flächen deutlich von den anderen beiden Systemen unterschieden und die höchste Heterogenität zeigten. Das bedeutet, dass mit der Einbringung von Buche eine Erhöhung der strukturellen Vielfalt zu erreichen ist, was als positiv zu werten ist, da sich die Artenvielfalt im Laufe der Zeit, allein schon wegen der höheren Verfügbarkeit differenzierter Nischen, erhöhen kann (Levine & Hille Ris Lambers 2009). Auch wirkt sich eine erhöhte strukturelle Heterogenität und ein größerer Baumartenreichtum günstig auf die Stabilität von Waldökosystemen aus (z. B. Jactel et al. 2017; Park et al. 2019).
Die strukturellen Veränderungen der Bestände sind dabei direkt bzw. indirekt auf die Bewirtschaftungsmaßnahmen zurückzuführen. So steht die größere Baumartenvielfalt im Altbestand bzw. der Verjüngung in unmittelbarem Zusammenhang mit den Bewirtschaftungsmaßnahmen ( → gezielte Einbringung der Buche). Die Effekte der Buchenbeimischung auf den Humuszustand oder das Mikrohabitatangebot sind dagegen eher indirekter Natur. So ist ein wesentlicher Effekt der Buchenbeimischung, dass es durch die – verglichen mit der Nadelstreu der Fichte – rascher zersetzbare Laubstreu der Buche zu einer sukzessiven Änderung des Humuszustandes kommt (z. B. Kudernatsch et al. 2021). Zu einem Wandel der Humusform kommt es dabei insbesondere in den laubholzdominierten Bestandesbereichen, weshalb Fichten-Buchen-Mischbestände eine höhere räumliche Variabilität der Humusauflage aufweisen.
Einen wichtigen Teil der strukturellen Vielfalt im Wald stellen Habitatbäume dar. Darunter werden lebende oder auch abgestorbene Bäume verstanden, die besondere Kleinlebensräume, sogenannte Mikrohabitate, aufweisen, wie z. B. Baumhöhlen, Stammverletzungen oder Pilzkonsolen. Unser Projekt konnte zeigen, dass die Beimischung von Buche in Fichtenreinbestände das Vorkommen derartiger Strukturen begünstigt, wobei insbesondere das Angebot an Kleinhöhlen erhöht wurde (Kudernatsch & Schauer 2022). Dies ist in Einklang mit anderen Untersuchungen, die zeigen konnten, dass Laubbäume durchschnittlich mehr Mikrohabitate ausbilden als Nadelbäume (Vuidot et al. 2011). Dabei sind insbesondere Höhlen häufiger an Laub als an Nadelbäumen anzutreffen (Larrieu et al. 2012).
Alpha-Diversität
Häufig wird postuliert, dass eine Erhöhung der Artenvielfalt in der Baumschicht einen »diversitätsfördernden Effekt« hat (z. B. Ellenberg & Leuschner 2010; Gamfeldt et al. 2013). Tatsächlich wurde im Rahmen unserer Untersuchungen allerdings kein klarer positiver Zusammenhang zwischen der Erhöhung der Baumartenvielfalt durch Waldumbau und der lokalen Artenvielfalt gefunden. So war über alle Artengruppen hinweg die Alpha-Diversität der H-Flächen nicht höher als die der Fichtenreinbestände. Gleiches gilt für die meisten der untersuchten Artengruppen.
Dass es bezüglich der Alpha-Diversität keinen einheitlichen, artengruppenübergreifenden Trend gibt, hängt u. a. damit zusammen, dass sich die betrachteten Artengruppen hinsichtlich ihrer Reaktion zum Teil unterscheiden. So wird die Pilzartenvielfalt im vorliegenden Fall beispielsweise durch den Waldumbau mit Buche begünstigt, während die lokale Diversität der Käfer eher einen negativen Trend zeigt. Dies bestätigt die Ergebnisse bisheriger Studien, die ebenfalls Unterschiede in der Reaktion zwischen verschiedenen Artengruppen aufzeigen konnten, wobei reine Nadelholzforste bzgl. einzelner Artengruppen mitunter durchaus artenreicher waren als Umbaubestände (z. B. Engel & Ammer 2001; Engelhard & Reif 2004; Budde et al. 2011). Als weitere Gründe für die begrenzte Reaktion der lokalen Artenvielfalt können das vergleichsweise niedrige Alter der Umbaubestände (das Maximalalter der Buchen in den Waldumbausystemen beträgt durchschnittlich 51 Jahre) sowie die Historie des Ebersberger Forstes genannt werden. Durch die intensive Nadelholzwirtschaft während der letzten zwei Jahrhunderte dürfte die Habitattradition vieler auf Laubholz bzw. Buche spezialisierter Arten unterbrochen worden sein, sodass eine Wiederbesiedlung aufgrund fehlender Spenderflächen unter Umständen lange Zeiträume benötigt. Dies gilt insbesondere für Arten bzw. Artengruppen, die durch eine eingeschränkte Mobilität gekennzeichnet sind, hingegen weniger für Arten, die kaum verbreitungslimitiert sind (wie z. B. die Pilze, deren zahlreiche Sporen weit fliegen können; Krah & Bässler 2021).
Beta-Diversität
Wesentlich deutlichere Unterschiede zwischen den drei Systemen zeigen sich, wenn man die Betadiversität betrachtet. Dieser Befund dürfte mit der Tatsache zusammenhängen, dass sich die Flächen der H-Variante bezüglich ihrer Strukturen und damit letztlich auch ihres Nischenangebots am deutlichsten voneinander unterscheiden, während sich die Fichtenreinbestände sowie die Flächen mit geringen Buchenanteilen untereinander tendenziell ähnlicher sind. So gab es z. B. hinsichtlich der Mischungsform klare Unterschiede zwischen den einzelnen Flächen der H-Variante. Während die Buchen auf manchen Flächen eher trupp- bis gruppenweise beigemischt waren, war für andere Flächen eine eher horst- bis flächenweise Beimischung charakteristisch. Merkliche Unterschiede zwischen den einzelnen Flächen der H-Variante gab es auch hinsichtlich des Kronenschlusses oder der vertikalen Differenzierung der Bestände. Insofern zeichnen sich die Flächen mit hohem Buchenanteil durch eine – verglichen mit dem Z- bzw. L-System – hohe Variabilität der Strukturen aus, wodurch auf den einzelnen Flächen verschiedenartige Nischen bzw. Nischenkombinationen entstehen, die von entsprechend unterschiedlichen Arten als Habitat genutzt werden können. Ein hoher »Arten-Turnover« zwischen den einzelnen Flächen und somit eine höhere Beta-Diversität sind die Folge.
Heterogene Artgemeinschaften sind aufgrund ihrer großen Bandbreite an ökologischen Funktionen ein wichtiger Teil der Funktionalität und Stabilität von Ökosystemen (Tilman 1996). Systeme mit artenreichen und heterogenen Artengemeinschaften weisen eine höhere Produktivität, verbesserte Nährstoffkreisläufe und eine höhere Resistenz gegenüber invasiven Arten oder Störungen auf (Cardinale et al. 2002; Hughes et al. 2004). Ein Grund hierfür ist, dass heterogenere Gemeinschaften den Wegfall bestimmter Arten nach großflächigen Störungen oder Änderungen der Umwelt besser kompensieren können (Allison 2004; Doak et al. 1998).
Gesamtartenvielfalt
Betrachtet man die im Rahmen unserer Studie ermittelten bzw. modellierten Gesamtartenzahlen, können kaum Unterschiede zwischen den drei Systemen aufgezeigt werden. Das Ersetzen aller Fichtenreinbestände durch ähnlich strukturierte Fichten-Buchen-Mischbestände würde entsprechend zu keinem deutlichen Anstieg der Gesamtartenvielfalt auf Landschaftsebene führen. Eine Zunahme der Gesamtdiversität auf Ebene der Landschaft kann hingegen erreicht werden, wenn zwei hinsichtlich ihrer Artengemeinschaften möglichst verschiedene (Z, H) bzw. alle drei Bewirtschaftungssysteme miteinander kombiniert werden. Durch die Kombination der verschiedenen Bewirtschaftungssysteme wird die Anzahl potenzieller Habitate und Nischen innerhalb der Landschaft erhöht. Dadurch können die Ansprüche verschiedenster Arten, von »Fichtenspezialisten« über »Generalisten« bis hin zu »Laubholz- bzw. Buchenwaldarten«, abgedeckt werden, was einen Anstieg der Gesamtdiversität zur Folge hat.
Die für den Ebersberger Forst durchgeführten Berechnungen bestätigen die bereits in anderen Studien getätigte Beobachtung, dass eine höhere Diversität der Betriebsformen in der Landschaft die Diversität positiv beeinflusst (Ammer et al. 2017; Schall et al. 2018). Heinrichs et al. (2020) konnten ferner zeigen, dass sich eine Mischung von Reinbeständen der beiden Baumarten Fichte und Buche gegenüber Baumartenmischungen innerhalb eines Bestandes positiv auf die Diversität auf Landschaftsebene auswirkt. Dies zeigt, dass rein unter Diversitätsgesichtspunkten somit auch der Erhalt eines gewissen Anteils an Reinbeständen durchaus sinnvoll sein kann.
Fazit und Handlungsempfehlungen
Zusammenfassend konnte gezeigt werden, dass der Umbau von Fichtenreinbeständen dazu beiträgt, die strukturelle Vielfalt, Diversität und damit Funktionalität der Wälder zu erhöhen. Da die positiven Effekte v. a. bei der Beta-Diversität sowie der strukturellen Vielfalt vielfach erst bei höheren Buchenanteilen auftraten, kann bei isolierter Betrachtung der drei Varianten das Bewirtschaftungssystem mit hohen Buchenanteilen als das ökologisch vorteilhafteste angesehen werden. Bezieht man allerdings die Ebene der Landschaft in die Bewertung mit ein, zeigt sich, dass insbesondere eine Kombination aller drei Systeme (bzw. der zwei Systeme, die sich am deutlichsten voneinander unterscheiden → Z, L) unter Diversitätsgesichtspunkten am besten abschneidet. Insofern erscheint es in von Fichten dominierten Landschaften ratsam, zusätzlich zu einem Umbausystem mit höheren Buchen-/Laubholzanteilen möglichst auch noch andere Bewirtschaftungsvarianten zu realisieren. Im Ebersberger Forst mit seiner Nähe zum Alpenrand und seinen vergleichsweise hohen Niederschlägen kann dabei auch der Erhalt einzelner Fichtenreinbestände eine ökologisch sinnvolle Option sein. Dadurch kann auch den an Nadelholz gebundenen Arten – zumindest mittelfristig – noch ein Auskommen ermöglicht werden.
Um eine große räumliche Heterogenität und damit Nischenvielfalt zu gewährleisten, sollte bei der Beimischung der Buche auf möglichst unterschiedliche Mischungsformen und -anteile geachtet werden, da dies einen hohen Artenwechsel zwischen den einzelnen Umbau-Beständen erwarten lässt. Auch der Faktor Licht spielt hinsichtlich der Ausbildung unterschiedlicher Artengemeinschaften eine große Rolle. Insofern sollte im Zuge des Waldumbaus gezielt darauf hingearbeitet werden, unterschiedliche Belichtungssituationen in den Beständen zu generieren, um sowohl Licht- als auch eher Schattenliebenden Arten ein Vorkommen zu ermöglichen. Auch gilt es das in den Umbaubeständen vorhandene große Potenzial an Mikrohabitaten zu fördern und für die Zukunft zu erhalten, wobei insbesondere auch den Pionier- und Weichlaubhölzern gezielt Beachtung geschenkt werden sollte.
Schließlich und letztendlich sollte die Totholzmenge und Vielfalt in den Beständen erhöht werden, indem Totholz möglichst verschiedener Baumarten belassen bzw. aktiv angereichert wird. Aufgrund der erhöhten Baumartenvielfalt bieten die Waldumbaubestände hierzu gute Voraussetzungen. Die Totholzanreicherung sollte dabei gleichermaßen in sonnigen wie in schattigen Bestandesbereichen erfolgen (vgl. Krah & Bässler 2021).
Summary
In the Ebersberger Forest, the largest enclosed wooded region in the lowlands of Southern Germany, we investigated the effects of forest conversion from pure spruce stands to mixed spruce-beech forests on structural heterogeneity and species diversity. Therefore, three management systems were compared (pure spruce stands vs. mixed spruce-beech stands with low and high proportions of beech). We showed that the admixture of beech is leading to an increase in structural and species diversity, resulting in a higher functionality of the forest stands. Our study suggests that the influence of the proportion of beech has an important impact especially on beta diversity, even to a higher degree than on alpha diversity and total species richness. Combining pure spruce and mixed spruce-beech stands at the landscape scale can help to increase landscape level biodiversity and conserve species adapted to conifer forests.
Literatur
- Allison, G. (2004): The influence of species diversity and stress intensity on community resistance and resilience. Ecological Monographs (74), S. 117–134
- Ammer, C.; Schall, P.; Goßner, M. M.; Heinrichs, S.; Boch, S.; Prati, D.; … Fischer, M. (2017): Waldbewirtschaftung und Biodiversität: Vielfalt ist gefragt. AFZ - Der Wald. (72), S. 20–25
- Böhme, J. (2001): Phytophage Käfer und ihre Wirtspflanzen in Mitteleuropa. Bioform, Heroldsberg
- Budde, S.; Schmidt, W.; Weckesser, M. (2011): Impact of the admixture of European beech (Fagus sylvatica L.) on plant species diversity and naturalness of conifer stands in Lower Saxony. Waldökologie, Landschaftsforschung und Naturschutz (11), S. 49–61
- Cardinale, B. J.; Palmer, M. A.; Collins, S. L. (2002): Species diversity enhances ecosystem functioning through interspecific faciliation. Nature (415), 426–429
- Doak, D. F.; Bigger, D.; Harding, E. K.; Marvier, M. A.; O'Malley, R. E.; Thomson, D. (1998): The statistical inevitability of stability-diversity relationships in community ecology. The American Naturalist, 151(3), S. 264–276
- Ellenberg, H.; Leuschner, C. (2010): Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht (6. Aufl.). Eugen Ulmer, Stuttgart
- Engel, K.; Ammer, U. (2001): Analyse und Bewertung von Umbaumaßnahmen in Fichtenbeständen anhand ökologischer Gilden der Wirbellosen-Fauna. Forstwirtschaftliches Centralblatt. (120), S. 242–255
- Engelhard, J.; Reif, A. (2004): Veränderungen der Bodenvegetation durch Fichtenanbau auf Standorten des Kalkbuchenwaldes. Waldökologie online (1), S. 29–56
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; … Bengtsson, J. (2013): Higher levels of multiple ecosystem services are found in forests with more tree species. Nature Communications, 4, 1340. https://doi.org/10.1038/ncomms2328
- Heinrichs, S.; Ammer, C.; Mund, M.; Boch, S.; Budde, S.; Fischer, M.; … Schall, P. (2019): Landscape-scale mixtures of tree species are more effective than stand-scale mixtures for biodiversity of vascular plants, bryophytes and lichens. Forests, 10(1), 73, https://doi.org/10.3390/f10010073
- Heinrichs, S.; Schall, P.; Ammer, C.; Fischer, M.; Goßner, M. M. (2020): Annahmen und Ergebnisse zur Biodiversität im Wirtschaftswald – Neues aus der Biodiversitätsforschung. WSL Berichte (100), S. 15–29
- Hughes, R. A.; Stachowicz, J. J. (2004): Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. PNAS, S. 8998–9002
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; … Brockerhoff, E. G. (2017): Tree diversity drives forest stand resistance to natural disturbances. Current Forestry Reports, 3(3), S. 223–243
- Jerz, H. (1993): Das Eiszeitalter in Bayern. Erdgeschichte - Gesteine - Boden. Schweizerbart, Stuttgart
- Krah, F. S.; Bässler, C. (2021): Totholzpilze – Baumart wichtiger als Totholzgröße und Besonnung. AFZ - Der Wald. (16/2021), S. 22–23
- Kudernatsch, T.; Schauer, B.; Walentowski, H. (2021): Effects of forest conversion of spruce monocultural stands to mixed spruce-beech forests on vegetation and humus layer. Tuexenia. (41), S. 109–132
- Kudernatsch, T.; Schauer, B. (2021): Mehr Vielfalt durch Waldumbau. LWF aktuell 1/2022, S. 15–17
- Larrieu, L.; Cabanettes, A.; Delarue, A. (2012): Impact of silviculture on dead wood and on the distribution and frequency of tree microhabitats in montane beech-fir forests of the Pyrenees. European Journal of Forest Research, 131(3), S. 773–786
- Levine, J. M.; Hille Ris Lambers, J. (2009): The importance of niches for the maintenance of species diversity. Nature, 461(7261), S. 254–257
- Park, J.; Kim, H. S.; Jo, H. K.; Jung, B., I. (2019): The influence of tree structural and species diversity on temperate forest productivity and stability in Korea. Forests, 10(12), 1113. https://doi.org/10.3390/f10121113
- PIK (2009): Klimadaten und Szenarien für Schutzgebiete. www.pik-potsdam.de/~wrobel/sg-klima-3/nav_bl.html, Stand:19.04.2022
- Schall, P.; Gossner, M. M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; … Mori, A. (2018): The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. Journal of Applied Ecology, 55(1), S. 267–278
- Sponholz, H. (1975): Ebersberger Forst, Geschichte und Gegenwart. Verlag H. Kliempt, Ottenhofen/Mkt. Schwaben
- Storch, F.; Dormann, C. F.; Bauhus, J. (2018): Quantifying forest structural diversity based on large-scale inventory data: A new approach to support biodiversity monitoring. Forest Ecosystems, 5(1), 230; https://doi.org/10.1186/s40663-018-0151-1
- Teuffel, K. von; Baumgarten, M.; Hanewinkel, M.; Konold, W.; Sauter, U. H.; Spiecker, H.; Wilpert, K. von (2005): Waldumbau für eine zukunftsorientierte Waldwirtschaft – Ergebnisse aus dem Südschwarzwald. Springer, Berlin, Heidelberg
- Tilman, D. (1996): Biodiversity: Population versus ecosystem stability. Ecology (77), S. 350–363
- Vuidot, A.; Paillet, Y.; Archaux, F.; Gosselin, F. (2011): Influence of tree characteristics and forest management on tree microhabitats. Biological Conservation, 144(1), S. 441–450
- Walentowski, H.; Gulder, H. J.; Kölling, C.; Ewald, J.; Türk, W. (2001): Die regionale natürliche Waldzusammensetzung Bayerns. LWF Wissen (32), 99 S.
Beitrag zum Ausdrucken
Weiterführende Informationen
Autoren



 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden
 Zoombild vorhanden
Zoombild vorhanden